

Research Article | DOI: https://doi.org/10.31579/2834-5010/007
Impaired Energy Metabolism and Altered Brain Histoarchitecture Characterized by Inhibition of Glycolysis and Mitochondrial Electron Transport-Linked Enzymes in Rats Exposed to Diisononyl Phthalate
1 Department of Environmental Health Science, Faculty of Basic Medical Sciences, Ajayi Crowther University, Oyo, Nigeria.
2 Biochemistry Program, Department of Chemical Sciences, Faculty of Natural Sciences, Ajayi Crowther University, Oyo, Nigeria.
3 Department of Biomedical Sciences, Faculty of Medicine and Health Science, Universiti Putra Malaysia. Malaysia.
4 Anatomy Unit, Faculty of Basic Medical Sciences, Ajayi Crowther University, Oyo, Nigeria.
*Corresponding Author: Samuel Abiodun Kehinde, Department of Environmental Health Science, Faculty of Basic Medical Sciences, Ajayi Crowther University, Oyo, Nigeria.
Citation: Samuel A Kehinde., Ore A., Abosede T Olajide., and Ebenezer O Ajiboye., (2022). Impaired Energy Metabolism and Altered Brain Histoarchitecture Characterized by Inhibition of Glycolysis and Mitochondrial Electron Transport-linked Enzymes in Rats Exposed to Diisononyl Phthalate. International Journal of Clinical Therapeutics. 2(3); DOI: 10.31579/2834-5010/007
Copyright: © 2023, Samuel Abiodun Kehinde. Nguyen, this is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 03 August 2023 | Accepted: 01 September 2023 | Published: 01 June 2023
Keywords: diisononyl phthalate; cellular respiration; glycolysis; mitochondrial electron transport complexes; oxidative phosphorylation; brain; rat
Abstract
The brain is an energy demanding organ, constituting about 20% of the body's resting metabolic rate. An efficient energy metabolism is critical to neuronal functions. Glucose serves as the primary essential energy source for the adult brain and plays a critical role in supporting neural growth and development. The potential for endocrine disrupting chemicals (EDCs) such as phthalates to have a negative impact on neurological functions has been widely recognized. We examined the impact of diisononyl phthalate (DiNP) on neural energy transduction by using cellular energy metabolizing enzymes as indicators. Over the course of 14 days, eighteen (18) albino rats divided into three groups (1,2 and 3) of six albino rats were given Tween-80/saline, 20 and 200 mg/kg body weight respectively. In the brain, we assessed histological changes as well as activities of selected enzymes of energy metabolism such as the glycolytic pathway, citric acid cycle and mitochondrial electron transport-linked complexes. Activities of the glycolytic enzymes assayed for were down-regulated, and the activities of the TCA cycle enzymes were significantly decreased (P<0.05) except citrate synthase activity with no statistically significant change following the administration of DiNP. Also, respiratory chain complexes (Complex I-IV) activities were significantly (P<0.05) down-regulated when compared to control. DiNP exposure altered the histological integrity of various brain sections. These include degenerated Purkinje neurons, distortion of the granular layer and Purkinje cell layer. Data from this study indicated impaired brain energy metabolism and altered brain histoarchitecture from DiNP exposure.
Introduction
Endocrine disrupting chemicals (EDCs) have been shown to have the potential to negatively affect neuro-development, and credible mechanisms have been postulated [1]. Since phthalates are a type of EDC, earlier reviews by Braun [1], Benjamin et al. [2], and Ejaredar et al. [3], have suggested a connection between phthalate exposure and a number of neuro-developmental outcomes, such as autism, lowered intelligence quotient (IQ), and impairment of both mental and psychomotor development. Due to the ubiquitous use of phthalates in consumer and commercial items, phthalate exposure in humans is prevalent, mostly through oral ingestion but also via inhalation and skin contact [4]. The propensity of phthalates to transverse the placenta brings up several questions regarding the developmental outcomes of in-utero exposure, especially when in combination with the potentiality for increased sensitivity in the unborn and child [1], according to Langonne et al. [5]. After exposure, phthalate diesters are quickly transformed into mono-esters and eliminated in the urine [4]. The half-lives of different phthalate metabolites are reported to be between 3 and 18 hours.
The results of a study by Polanska et al. [6] show that exposure to phthalates during pregnancy may be detrimental to an infant's neuro-development, highlighting the need for regulations and public health initiatives targeted at limiting this exposure. Furthermore, it is interesting that prenatal phthalate exposure's neurotoxicity has been extensively studied in animal research [7]. While some studies have revealed that some phthalates are detrimental to children's neuro-development [8, 9], others have found no significant association, a sex-specific effect, or a positive effect [10; 11]. The hypothalamic-pituitary-gonadal, adrenal, and thyroid axes are all dysregulated by phthalates, which is crucial for neuro-development, according to a systematic review conducted by Hlisnková et al. [12]. Phthalates interfere with nuclear receptors at the intracellular level in numerous neuronal structures, affecting brain functions and the development of neurological disorders.
Diisononyl phthalate (DiNP) takes the place of other common phthalates. It can be found in a wide range of things, such as toys, consumer goods, food packaging, and building materials. DiNP is readily capable to pass through saliva and be eaten since it is not covalently bonded to polymers [13]. DiNP can enter the body through ingestion, inhalation, or skin absorption. The European Chemicals Bureau estimated that children are exposed to 200 g/kg/day of DiNP through toys [13]. Currently, there are limited or no study that have directly examined the role of exposure to DiNP on activities of neuronal energy metabolizing enzymes. Hence, this study utilized cellular energy metabolizing enzymes as biomarker to investigate the impact of DiNP on neural energy transduction.
Materials and Methods
2.1 Chemicals and reagents
CYPRESS® Diagnostics from Belgium supplied the lactate dehydrogenase assay kit and we sourced DiNP from Relonchem Ltd. UK. We ensured the highest analytical quality for all additional reagents, chemicals and kits used.
2.2 Animals
Eighteen (18) albino rats, weighing between 200-230g, were procured from the animal breeding facility of the University of Ibadan College of Medicine, Ibadan. Subsequently, the rats were acclimated and subjected to treatment while being housed in plastic cages at the animal facility of the Department of Chemical Sciences, Ajayi Crowther University. Throughout the study, the rats had ad libitum access to water and pelletized food. Approval by the Faculty Committee on Animal Ethics for the use of these animals was granted with authorization number FNS/ERC/2021/006.
2.3 Experimental groups
The rats were stratified into three cohorts, each consist of six rats. Group A, the control group, received Tween-80. Group B received a dosage of 20 mg/kg/BW of DiNP, while Group C was administered 200 mg/kg of DiNP orally (Fig. 1). The exposure lasted 14 days. The DiNP solution was prepared using a mixture of normal saline and Tween-80 in a 1:1 v/v ratio. The selection of the doses (20 and 200 mg/kg/day DiNP) was based on previous research findings [15, 16].
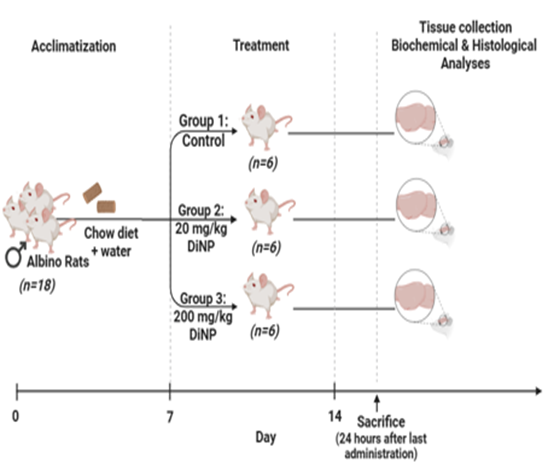
Figure 1: Experimental protocol (created in www.biorender.com)
2.4 Organs sampling and biochemical techniques
We adhered to guidelines established for the ethical management and care of laboratory animals during the handling of the rats [17]. Following the final dosage, the animals were euthanized, and the brain was excised while they were under diethyl ether anesthesia. Subsequently, the extracted brain was rinsed in pre-cooled KCL (1.15%), blotted dry, weighed, and afterwards homogenized using 10 volumes per weight (v/w) of phosphate buffer (0.1 M ; pH 7.4) in a potter-elevehjem homogenizer. The resultant homogenate was then centrifuged (10,000 g; 15 minutes; 4°C) to isolate the supernatant for subsequent biochemical assays.
To process the rats’ brains and isolate the mitochondria, we followed the methodology previously outlined by Erika et al. [18]. The determination of hexokinase (HK) activity followed the protocol described by Colowick [19], while the Phosphofructokinase (PFK) activity was assessed following an established method of Sims and Blass [20]. Aldolase (ALD) activity measurement adhered to the procedure outlined by Jagannathan et al. [21]. The manufacturer's instructions for the LDH Kit (LABKIT) were followed to determine the lactate dehydrogenase (LDH) activity. Lastly, the determination of NADase activity followed the protocol outlined by Tatsuno et al. [22].
We employed the spectrophotometric enzyme assay method as outlined by Yu et al. [23] to evaluate citrate synthase activity. Using a method previously published by Romkina and Kiriukhin [24], the activity of isocitrate dehydrogenase (IDH) was assessed spectrophotometrically at an absorbance of 340 nm by quantifying the inhibition in NAD+. Alpha-ketoglutarate (α-KGD) activity was assessed using the α-KGDH Kit in accordance with the supplier's recommendations from BioVision Incorporated. To determine Malate Dehydrogenase activity, we followed the protocol previously outlined by López-Calcagno et al. [25].
To evaluate mitochondrial function, we quantified the activities of electron transport-linked enzymes (Complex I; NADH ubiquinone oxidoreductase, Complex II; succinate ubiquinone oxidoreductase, Complex III; cytochrome c oxidoreductase, and Complex IV; Cytochrome C Oxidase) in the mitochondria, following the methodology outlined by Medja et al. [26].
Furthermore, we quantified the neural total protein concentration utilizing the Biuret protocol as outlined by Gornall et al. [27]. This method forms a chelate (purple-colored) between Cu2+ ions and the protein’s peptide bonds which has a maximum absorbance at 540 nm.
2.5 Histopathological analysis of brain tissue
The fixed brain slices were dehydrated (using ethanol and xylene) and paraffin-embedded. Subsequently, hematoxylin and eosin (H&E) was used to stain them and processed for histological analysis using a Leica DM750 camera microscope.
2.6 Statistical analysis
Results are shown as the mean ± standard deviation (SD). Analysis of variance (ANOVA) was used to evaluate the homogeneity of the group. Where heterogeneity was observed, Tukey's test was to distinguish between the groups. P-values < 0>
Results
3.1 Activities of neural glycolytic enzymes in rats exposed to DiNP.
As depicted in Fig. 2, exposure to DiNP resulted in significant down-regulations (P < 0>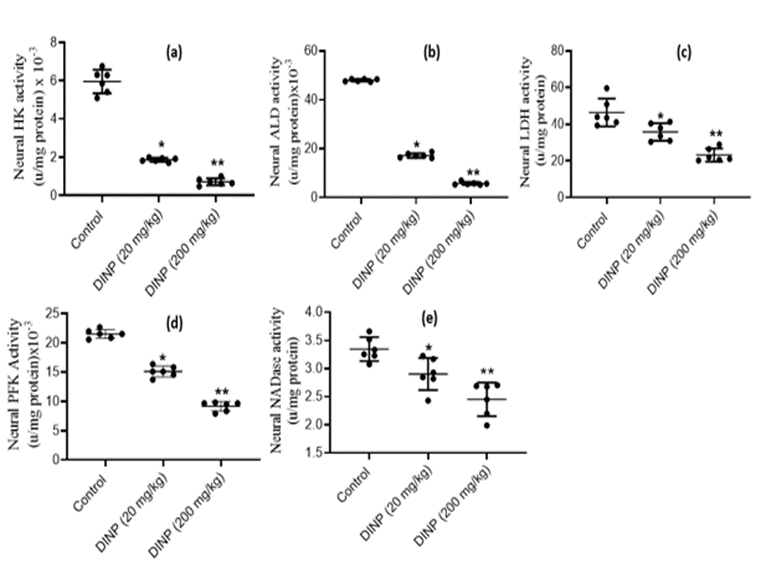
Figure 2: Effect of DiNP on activities of neural glycolytic enzyme in rats: (a) hexokinase - HK, (b) aldolase - ALD, (c) lactate dehydrogenase - LDH, (d) phosphofructokinase - PFK, (e) NAD-nucleosidase - NADase. The data is represented as Mean ± Standard Deviation (SD) for each group of six rats. * denotes statistical significance (P < 0>
3.2 Activities of neural tricyclic acid cycle enzymes in rats exposed to DiNP.
Fig. 3(A–D) illustrates the impact of DiNP on the activity of enzymes of tricarboxylic acid cycle in rat brain. Rats administered DiNP (20 and 200 mg/kg) did not display any discernible change in brain citrate synthase activity, which was statistically insignificant relative to the control group. However, notable down-regulations (p < 0>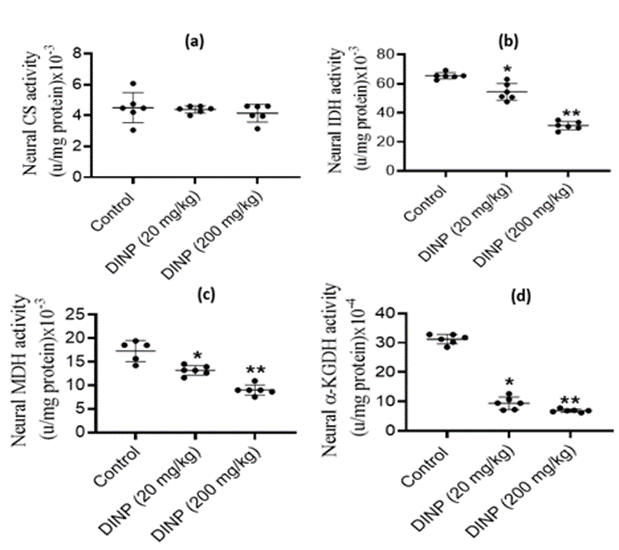
Figure 3: Effect of DiNP on neural tricyclic acid cycle enzymes activities in rats: (a) citrate synthase - CS, (b) isocitrate dehydrogenase - IDH, (c) malate dehydrogenase - MDH, (d) alpha ketoglutarate dehydrogenase - α-KGDH. The data is presented as Mean ± Standard Deviation (SD) for each group of six rats: (a) * denotes statistical significance (P < 0>
3.3 Activities of neural electron transport chain complexes in rats exposed to DiNP.
The activity levels of brain electron transport chain enzymes exhibited significant (P < 0>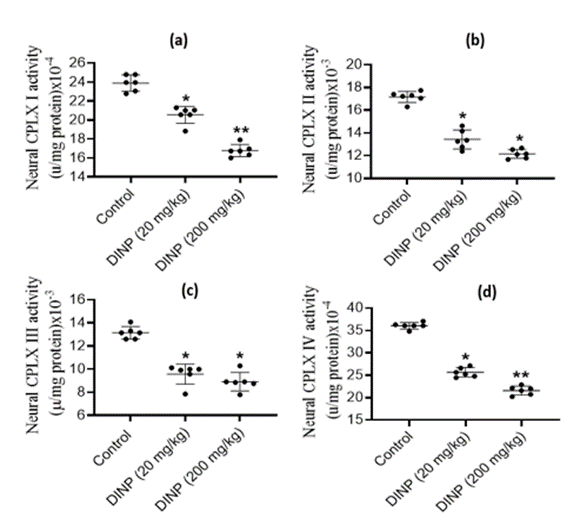
Figure 4: Effect of DiNP on neural electron transport chain complexes activities in rats: (a) complex I - CPLX 1, (b) complex II - CPLX II, (c) complex III - CPLXIII, (d) complex IV - CPLXIV. The data is presented as Mean ± Standard Deviation (SD) for each group of six rats. * Denotes statistical significance (P < 0>
3.4 Histopathological analysis of brain tissue
The histoarchitecture of the brain tissue from rats exposed to DiNP (20 and 200mg/kg) is shown in Fig. 5, 6, and 7, along with a score graph that depicts the degree of the damage. With a high dose of DiNP (200 mg/kg), neuropil distortion is shown. The neuropil appears to be fragmented, as shown in plates 2 and 3. Pyramidal neurons displayed pyknotic behavior and appeared smaller and darker. Comparing groups A (Control) and B (20mg/kg) to group C (200mg/kg), undamaged neurons can be found in
much higher numbers in the various levels of the cerebellum. This is a warning sign for the neuro-degeneration shown at 200 mg/kg. At 200mg/kg, the granule neurons seemed to have shrunk. Both Groups A and B appeared to have intact neuronal morphology. In the nucleolus of the undamaged neurons, the nissl material was plainly visible. Relative to the control group and 20mg/kg DiNP, protein integrity was clearly visible, demonstrating the viability of the neurons' nissl substances.
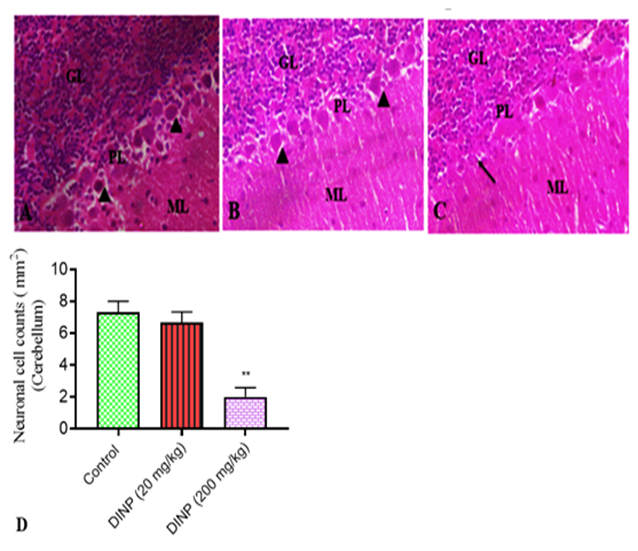
Figure 5: Representative sections of the cerebellum stained with hematoxylin and eosin from rats exposed to DiNP.
A connotes control group, B - 20 mg/kg DiNP group, C-200 mg/kg DiNP group, and D- neuronal cell counts. Notable regions include the molecular layer - ML, Purkinje cell layer - PCL, and granular layer - GL. Normal Purkinje neurons are marked with arrowheads. Arrows indicates degenerated purkinje neurons. * Denotes statistical significance (P < 0>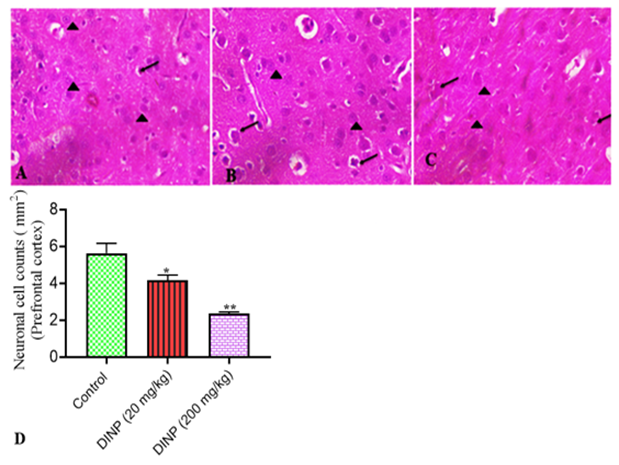
Figure 6: Representative sections of the prefrontal cortex stained with hematoxylin and eosin from rats exposed to DiNP.
A connotes control group, B- 20 mg/kg DiNP group, C - 200 mg/kg DiNP group, and D- neuronal cell counts. Highlighted are dark pyramidal neurons (arrows) and normal pyramidal neurons (arrowheads). * Denotes statistical significance (P < 0>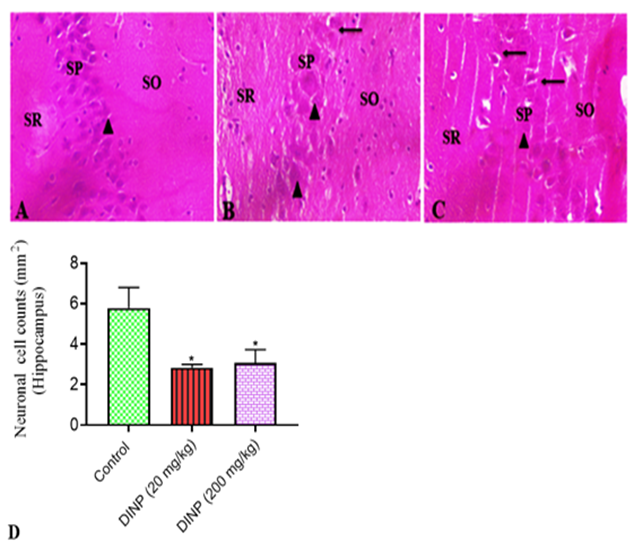
Figure 7: Representative sections of the hippocampus stained with hematoxylin and eosin from rats exposed to DiNP.
A connotes control group, B - 20 mg/kg DiNP group, C- 200 mg/kg DiNP group, and D- neuronal cell counts. Regions of interest include the stratum oriens - SO, stratum pyramidalis - SP, and stratum radiatum - SR. Arrowhead indicated normal pyramidal neurons. Arrows connotes dark pyramidal neurons. * denotes statistical significance (P < 0>
Discussion
Despite making up only 2% of total body weight, the brain is an "expensive" organ that contributes about 20% to resting metabolic rate of the body. The main required energy source for the adult brain is glucose, which is also crucial for the developing brain. Locally, rate of blood flow (to remove waste products and supply fuel), metabolic demand, and brain activity are all intimately connected. The blood flow rate to neural tissue increases with increased cellular activity associated with the particular brain function (e.g. processing information as a result of sensory stimulation or mental computations), which also increases the local demand for ATP [28]. Reduced activity, on the other hand, reduces blood flow and glucose and oxygen consumption. Both oxygen and glucose, which are essential for the growth and maintenance of the brain in both children and adults, must be continuously supplied to the brain. Transporters in endothelial cells, whose close connections make up the blood-brain barrier, take up substances utilized by the brain from the circulation [29].
Specialized transporters located in endothelial cells, which create the tight junctions of the blood-brain barrier, transport substances that cross the barrier from the bloodstream[29]. Examples of these transporters include the monocarboxylic acid transporter (MCT) and the glucose transporter (GLUT) [30]. Notably, the brain exhibits a high reliance on ketone bodies for energy utilization during the suckling period, as well as for serving as a carbon source in lipids, amino acids, and proteins synthesis, in contrast to pyruvate dehydrogenase and certain oxidative enzymes that display lower activity [30]. After birth, glycolytic and oxidative enzyme activity steadily increases with maturation and reaches adult levels soon after weaning [32]. An increase in oxidative glucose utilization has been linked to the proliferation of transporters on the blood-brain barrier [33]. These changes are also in line with gradually rising levels of enzymes involved in oxidative metabolism in the brain as well as rising activity of malate-aspartate shuttle enzymes [34]. As the brain develops, it consumes more glucose for energy, which is then converted into more pyruvate via the TCA cycle. This study provides the initial insights into the inhibitory impact of DiNP on enzymes associated with the glycolysis and oxidative phosphorylation.
The first critical point in controlling the pace of glucose metabolism in the brain is the glucose phosphorylation by hexokinase, which is considered an irreversible process; phosphofructokinase is the second important regulatory enzyme [35]. The brain commits glucose for use in the first stage of glycolysis. According to the results of this study, the experimental animals' exposure to 20 and 200 mg/kg DiNP considerably reduced their hexokinase activity compared to the control group. This demonstrates how DiNP inhibits brain glucose uptake by lowering hexokinase activity, which prevents glucose from being readily available for the glycolytic pathway. Fructose-6-phosphate and Mg-ATP are also the main and most intricate sources of energy for Phosphofructokinase-1, the main regulatory enzyme in responsible for regulating glycolysis [35]. It is basically irreversible, similar to other regulatory reactions. The delivery of DiNP significantly decreased PFK-1 activity, according to this experiment. Aldolase, a crucial glycolytic enzyme, plays a pivotal role in cleaving fructose 1-6-diphosphate to glyceraldehyde 3-phosphate and dihydroxy-acetone phosphate within the glycolytic metabolic pathway [36]. Previous studies have shown that Di[2-ethylhexyl] phthalate, a type of phthalate, did not exhibit any inhibitory effects on aldolase activity [37]. In contrast, our current study reveals that DiNP dose-dependently suppresses aldolase activity. It is important to note that the activity of PFK-1, an enzyme responsible for converting fructose 6-phosphate into fructose 1,6-bisphosphate (a required substrate for aldolase), has been down-regulated. Consequently, the readily available substrate for aldolase which is fructose-1-6-diphosphate, has decreased [38].
Furthermore, within the glycolytic pathway, the cytoplasmic NADH generated through the activity of glyceraldehyde-3-P dehydrogenase undergoes oxidation facilitated by LDH (a bidirectional enzyme). This LDH-mediated process is essential for the seamless progression of glycolysis [16]. The malate-aspartate shuttle activity is also indispensable to sustain the conversion of lactate into pyruvate, along with the generation of NADH + H, which subsequently serves as a vital oxidative fuel source. When O2 becomes scarce, glycolytic ATP synthesis can continue and even speed up since LDH in the brain functions in aerobic, hypoxic (low oxygen), anaerobic (zero oxygen), and ischemic (no blood flow) situations [32]. The amount of glucose that is converted to pyruvate or lactate in the presence of "sufficient" oxygen and without oxygen, respectively, is referred to as aerobic or anaerobic glycolysis. When the rate of aerobic glycolytic flux surpasses the capacity for pyruvate oxidation or the transport of NADH equivalents to the mitochondria, lactate production occurs. Since the utilization of lactate relies on its availability and transport through equilibrium mechanisms, it is considered an 'opportunistic' energy source. In cultured astrocytes and neurons, lactate is quickly taken in and oxidized, and oligodendroglia cells can use only a little of it for energy and lipid synthesis. [39]. A small amount of lactate serves as an energy source and contributes to lipid synthesis in oligodendroglia cells, while cultured astrocytes and neurons readily take up and oxidize it [39]. However, the results of this study indicate that DiNP, administered at both 20 and 200 mg/kg doses, significantly decreased LDH activity compared to the control group, thereby reducing the availability of this opportunistic energy source in the brain.
NADase, belonging to the hydrolase enzyme family, is responsible for hydrolyzing N-glycosyl compounds. It plays a role in nicotinate and nicotinamide metabolism, as well as in the calcium signaling pathway [40]. In cellular oxidation-reduction (redox) reactions, NAD+ and flavin adenine dinucleotide (FAD+) are crucial in capturing high-energy electrons and delivering them to the ETC to produce adenosine triphosphate (ATP). [41]. Numerous metabolic processes such as including fatty acid oxidation, glycolysis, the Krebs cycle, rely on NAD+ as a cofactor for various enzymes [42]. Given that NAD+ is an essential cofactor for enzymes involved in both glycolysis and TCA/ETC processes, the reduced NADase activity resulting from DiNP administration at doses of 20 and 200 mg/kg may impede these pathways. Consequently, this limitation in energy supply for brain functions could potentially disrupt these essential metabolic processes
The Krebs cycle, also recognized as the citric acid cycle, plays a fundamental role in glucose metabolism by transferring reducing equivalents to NADH and FADH2. These compounds collaborate with ETC to generate ATP and regenerate oxidized redox molecules. The TCA cycle accounts for the majority of ATP production from glucose metabolism. The initiation of oxidative metabolism occurs when pyruvate, derived from glycolysis, enters the mitochondrion and transforms into acetyl CoA. The TCA cycle is indispensable for energy generation, amino acid synthesis, and neurotransmission [43]. It commences with the transfer of a two-carbon acetyl group from Acetyl-CoA to a four-carbon acceptor molecule called oxaloacetate, facilitated by citrate synthase. This reaction leads to the formation of a six-carbon compound referred to as citrate. Importantly, the brain, like other tissues, hosts two isoforms of isocitrate dehydrogenase (IDH): the cytoplasmic NADP+-dependent IDH and the mitochondrial NAD+ - dependent IDH. The NAD-linked enzyme is allosterically regulated, with ATP and NADH inhibiting it, while ADP activates it. It catalyzes an essentially irreversible process involving the dehydrogenation of isocitrate to form oxalosuccinate [44]. Citrate synthase activity in our study remained unaffected by DiNP exposure (20 and 200 mg/kg), indicating the availability of the initial substrate required for the TCA cycle. However, IDH was found to be inhibited following DiNP administration.
The decreased activity of IDH as a result of DiNP exposure will diminish the cycle's role in energy production, resulting in reduced energy delivery to the brain. Malate dehydrogenase (MDH) catalyzes the reaction that converts of malate to oxaloacetate in the Krebs cycle. This reaction involves the oxidation of the hydroxyl group on malate and the reduction of NAD+. The cytosolic isoform of MDH contributes to the ETC as an electron donor, facilitating ATP production [45]. MDH exists in both mitochondrial and cytosolic isoforms. Alpha-ketoglutarate (α-KG) exerts significant influence over cellular energy utilization. It participates in multiple metabolic pathways, including its synthesis and breakdown. At a pivotal control point in the TCA cycle, α-KG undergoes decarboxylation by α-KG dehydrogenase to form succinyl-CoA and CO2. It is well-established that astrocytes convert glutamate into alpha-ketoglutarate to supply lactate, primarily derived from the Krebs cycle, to neurons [46]. Our investigation revealed that DiNP dramatically reduced MDH activity and the level of α-KG in a dose-dependent manner
Reducing equivalents, carried by molecules like NADH and FADH2, are transferred to the ETC either at complex I or II, while H+ ions are expelled across inner mitochondrial membrane. This process establishes a proton gradient that leads to the production of ATP. Notably, when protons enter the ETC through complex II, as opposed to CPLX I, fewer protons are ejected, resulting in a reduced ATP output. This method directly connects energy production to energy consumption since biological processes ultimately use ATP to produce ADP, the necessary precursor for the ETC [16].
The ETC comprises Complexes I- IV. The tricarboxylic acid (TCA) cycle generates a proton-motive force by utilizing electrons from NAD+ and FAD-linked enzymes, which is subsequently employed for ATP production. Specifically, electrons originating from FAD-linked SDH move from CPLX II to CPLX III through ubiquinone, while electrons from CPLX I directly enter CPLX III [47, 48]. This study represents the first investigation into the down-regulatory effects of DiNP on neural CPLX I, II, III, and IV of the ETC.
The results indicate that DiNP led to dose-dependent reductions in the activities of each complex. Prior research has shown that exposure to DEHP, a phthalate, resulted in decreased SDH activity [49]. Similarly, environmental pollutants such as benzyl butyl phthalate (TDI) and diphenylmethane diisocyanate (MDI) have been associated with reduced SDH activity and alterations in energy metabolism [50, 51]. The ETC uses NADH and FADH2, which are produced during glycolysis, oxidation, and other catabolic processes, to produce ATP. The down-regulated activities of the ETC complexes observed in rats exposed to DiNP demonstrate the inability of the brain to convert products of the glycolytic and TCA cycle into ATP. This explains why the ETC complexes were down-regulated.
The fact that neurons heavily rely on oxidative energy metabolism has suggested a unified histology mechanism of neuron degeneration based on studies of the underlying defective mitochondrial energy metabolism. It is not surprising that many mitochondrial disorders are encepha-cardiomyopathies given that the brain, heart, and skeletal muscles are among the body parts most dependent on energy [52, 53].
The reported results of DiNP inducing the pyramidal neurons to be pyknotic and shrunken, which is indicative of neuronal degeneration, reflect the emphasis given to mitochondrial failure as an executor in neural degeneration. In addition to the initial loss of ATP caused by the DiNP insult, the mitochondria's inability to produce the required energy has catastrophic effects on downstream systems like brain functioning and signaling. It is evident that a variety of neurodegenerative illnesses have impaired or inhibited mitochondrial energy metabolism as a major pathogenic component.
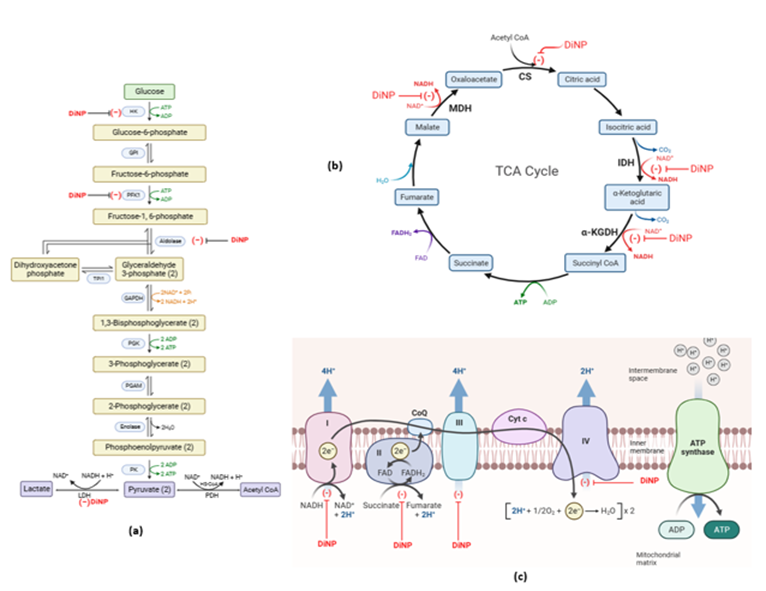
Figure 8: Mechanistic overview of the impact of DiNP on (a) neural glycolytic pathway, (b) neural citric acid cycle and (c) neural mitochondrial electron transport chain. (created at www.Biorender.com) .
Conclusions
It has been established that exposure to DiNP, a class of phthalates, is an endocrine disrupting substance. Data from this study showed that DiNP down-regulates neural glycolytic and mitochondrial metabolizing enzymes, which greatly perturbs the activities of enzymes involved in energy transduction in the brain.
Highlights
- Diisononyl phthalate (DiNP) downregulate the activities of neural glycolytic enzymes in rat
- DiNP caused significant decrease in activities of tricarboxylic acid cycle enzymes
- DiNP also downregulate the activities of electron transport complexes
- DiNP also caused alteration in brain histoarchitecture
Author Contributions
Conceptualization, S.A.K. and A.O.; methodology, S.A.K., A.O.; A.T.O. and E.A.O.; formal analysis, S.A.K., A.O.; A.T.O. and E.A.O.; investigation, S.A.K., A.O.; A.T.O. and E.A.O.; resources, S.A.K., A.O. and A.T.O.; data curation, S.A.K., A.O.; A.T.O. and E.A.O.; writing—original draft preparation, S.A.K.; writing—review and editing, S.A.K., A.O. and A.T.O.; supervision, S.A.K. and A.O.; project administration, S.A.K. and A.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The Institutional Review Board (or Ethics Committee) of the Faculty of Natural Sciences at Ajayi Crowther University gave its approval to the animal study protocol (protocol code FNS/ERC/2021/006).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors upon reasonable request.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest
References
- Braun JM (2017). Early-life exposure to EDCs: role in childhood obesity and neurodevelopment. Nature Reviews Endocrinology, 13(3): 161-173.
View at Publisher | View at Google Scholar - Benjamin S, Masai E, Kamimura N, Takahashi K, Anderson RC, et al (2017). Phthalate’s impact human health: Epidemiological evidences and plausible mechanism of action. Journal of hazardous materials, 340: 360-383.
View at Publisher | View at Google Scholar - Ejaredar M, Nyanza EC, Ten Eycke K, Dewey D (2015). Phthalate exposure and childrens neurodevelopment: A systematic review. Environ. Res, 142: 51–60.
View at Publisher | View at Google Scholar - Johns LE, Cooper GS, Galizia A, Meeker JD (2015). Exposure assessment issues in epidemiology studies of phthalates. Environ. Int. 85: 27–39.
View at Publisher | View at Google Scholar - Langonne L, Saillenfait AM, Payan JP (1998). Comparative embryotoxicity of di-nbutyl phthalate and its main metabolite mono-n-butyl phthalate at midgestations. Teratology 58: 22A.
View at Publisher | View at Google Scholar - Polanska K, Ligocka D, Sobala W, Hanke W (2014). Phthalate exposure and child development: the polish mother and child cohort study. Early. Hum. Dev, 90(9): 477–485.
View at Publisher | View at Google Scholar - Miodovnik A, Engel SM, Zhu C, Ye X, Soorya LV, et al. (2011). Endocrine disruptors and childhood social impairment. Neurotoxicolog 32(2): 261–267.
View at Publisher | View at Google Scholar - Engel SM, Miodovnik A, Canfield RL, Zhu C, Silva MJ, Calafat AM, et al. (2010). Prenatal phthalate exposure is associated with childhood behavior and executive functioning. Environ. Health. Perspect. 118(4):565–571.
View at Publisher | View at Google Scholar - Kim Y, Ha EH, Kim EJ, Park H, Ha M, et al, (2011). Prenatal exposure to phthalates and infant development at 6 months: prospective mothers and children's environmental health (moceh) study. Environ. Health. Perspect. 119(10):1495–1500.
View at Publisher | View at Google Scholar - Engel SM, Zhu C, Berkowitz GS, Calafat AM, Silva MJ, et al. (2019). Prenatal phthalate exposure and performance on the neonatal behavioral assessment scale in a multiethnic birth cohort. Neurotoxicology 30(4):522–528.
View at Publisher | View at Google Scholar - Whyatt RM, Liu XH, Rauh VA, Calafat AM, Just AC, et al. (2012). Maternal prenatal urinary phthalate metabolite concentrations and child mental, psychomotor, and behavioral development at 3 years of age. Environ. Health. Perspect. 120(2): 290–295.
View at Publisher | View at Google Scholar - Hlisníková H, Petrovičová I, Kolena B, Šidlovská M, Sirotkin A, et al. (2021) Effects and mechanisms of phthalates' action on neurological processes and neural health: a literature review. Pharmacol Rep. 73(2):386-404.
View at Publisher | View at Google Scholar - Silva MJ, Reidy JA, Preau Jr JL, Needham LL, Calafat AM et al. (2006). Oxidative metabolites of diisononyl phthalate as biomarkers for human exposure assessment. Environmental health perspectives. 114(8):1158-61.
View at Publisher | View at Google Scholar - Kransler KM, Bachman AN, McKee RH, (2012). A comprehensive review of intake estimates of di-isononyl phthalate (DiNP) based on indirect exposure models and urinary biomonitoring data. Regulatory Toxicology and Pharmacology. 62(2):248-56.
View at Publisher | View at Google Scholar - Ore A, Adewale AA, Kehinde SA, Faniyi TO, Oladeji AD, et al. (2022). Potential roles of oxidative stress and insulin resistance in diisononyl phthalate induced dyslipidemia and hepatosteatosis in BALB/c mice. Advances in Redox Research. 5:100038.
View at Publisher | View at Google Scholar - Kehinde SA, Ore A, Olajide AT, Ajagunna IE, Oloyede FA, et al. (2022). Diisononyl phthalate inhibits cardiac glycolysis and oxidative phosphorylation by down-regulating cytosolic and mitochondrial energy metabolizing enzymes in murine models. Advances in Redox Research. 2022; 100041.
View at Publisher | View at Google Scholar - NRC. Guide for the Care and Use of Laboratory Animals (1996). National academy press, Washington, D.C
View at Publisher | View at Google Scholar - Fernández-Vizarra E, Ferrín G, Pérez-Martos A, Fernández-Silva P, Zeviani M, et al. (2010). Isolation of mitochondria for biogenetical studies: An update. Mitochondrion. 10(3):253-62.
View at Publisher | View at Google Scholar - Colowick SP (1973) The Hexokinases. In The enzymes. 9:1-48.
View at Publisher | View at Google Scholar - Sims NR, Blass JP, (1986). Phosphofructokinase activity in fibroblasts from patients with Alzheimer's disease and age-and sex-matched controls. Metabolic Brain Disease. 1(1):83-90.
View at Publisher | View at Google Scholar - Jagannathan V, Singh K, Damodaran M, (1956). Carbohydrate metabolism in citric acid fermentation. 4. Purification and properties of aldolase from Aspergillus niger. Biochemical Journal. 63(1):94.
View at Publisher | View at Google Scholar - Tatsuno I, Sawai J, Okamoto A, Matsumoto M, Minami M, et al. (2007). Characterization of the NAD-glycohydrolase in streptococcal strains. Microbiology. 153(12):4253-60.
View at Publisher | View at Google Scholar - Yu X, Tesiram YA, Towner RA, Abbott A, Patterson E, et al. (2007) Early myocardial dysfunction in streptozotocin-induced diabetic mice: a study using in vivo magnetic resonance imaging (MRI). Cardiovascular diabetology. 6(1):1-8.
View at Publisher | View at Google Scholar - Romkina AY, Kiriukhin MY, (2017). Biochemical and molecular characterization of the isocitrate dehydrogenase with dual coenzyme specificity from the obligate methylotroph Methylobacillus Flagellatus. Plos one, 12: e0176056,
View at Publisher | View at Google Scholar - López-Calcagno PE, Moreno J, Cedeño L, Labrador L, Concepción JL, et al. (2009). Cloning, expression and biochemical characterization of mitochondrial and cytosolic malate dehydrogenase from Phytophthora infestans. Mycological research. 113(6-7):771-81.
View at Publisher | View at Google Scholar - Medja F, Allouche S, Frachon P, Jardel C, Malgat M, et al. (2009). Development and implementation of standardized respiratory chain spectrophotometric assays for clinical diagnosis. Mitochondrion. 9(5):331-9.
View at Publisher | View at Google Scholar - Gornall AG, Bardawill CJ, David MM. (1949). Determination of serum proteins by means of the biuret reaction. J. biol. Chem. 177(2):751-66.
View at Publisher | View at Google Scholar - Clarke DD, Sokoloff L. (1999). Circulation and energy metabolism. In G. J. Siegel, B. W. Agranoff, D. S. Albers, S. K. Fisher & M. D. Uhler (Eds.), Basic Neurochemistry (6th ed.). Philadelphia: Lippincott Williams and Wilkins.
View at Publisher | View at Google Scholar - Simpson IA, Dwyer D, Malide D, Moley KH, Travis A, et al. (2008). The facilitative glucose transporter GLUT3: 20 years of distinction. American Journal of Physiology-Endocrinology and Metabolism. 295(2): E242-53.
View at Publisher | View at Google Scholar - Nehlig A (2004) Brain uptake and metabolism of ketone bodies in animal models. Prostaglandins Leukot Essent Fatty Acids. 70:265–275.
View at Publisher | View at Google Scholar - Patel AB, Chowdhury GM, de Graaf RA, Rothman DL, Shulman RG, et al. (2005). Cerebral pyruvate car-boxylase flux is unaltered during bicuculline-seizures. Journal of Neuroscience Research, 79: 128–138.
View at Publisher | View at Google Scholar - McKenna MC, Gruetter R, Sonnewald U, Waagepetersen HS, Schousboe, A et al. (2006). Energy metabolism of the brain. In A., Siegel, G. J., Albers, R. W., Brady, S. T., Price, D. L., (Eds), Basic neurochemistry, 7th edition. Burlington, MA: Elsevier Academic Press. 531–557.
View at Publisher | View at Google Scholar - Vannucci SJ, Simpson IA. (2003). Developmental switch in brain nutrient transporter expression in the rat. American Journal of Physiology Endocrinology Metabolism, 285(5): E1127–E1134.
View at Publisher | View at Google Scholar - Ramos M, del Arco A, Pardo B, Martinez-Serrano A, Martinez Morales JR, et al. (2003). Developmental changes in the Ca2+-regulated mitochondrial aspartate– glutamate carrier aralar1 in brain and prominent expression in the spinal cord. Brain Research. Developmental Brain Research, 143(1): 33–46.
View at Publisher | View at Google Scholar - Keith Tornheim. (2018). Glucose Metabolism and Hormonal Regulation☆, Editor(s): Ilpo Huhtaniemi, Luciano Martini, Encyclopedia of Endocrine Diseases (Second Edition), Academic Press, pp :87-94.
View at Publisher | View at Google Scholar - Du S, Guan Z, Hao L, Song Y, Wang L, et al. (2014). Fructose-Bisphosphate Aldolase a Is a Potential Metastasis-Associated Marker of Lung Squamous Cell Carcinoma and Promotes Lung Cell Tumorigenesis and Migration. PLoS ONE. 9(1): e85804.
View at Publisher | View at Google Scholar - Mushtaq M, Srivastava SP, Seth PK. (1980). Effect of di-2-ethylhexyl phthalate (DEHP) on glycogen metabolism in rat liver. Toxicology, 16(2):153-161.
View at Publisher | View at Google Scholar - Kehinde SA, Ore A, Olajide AT, Adeyemo OA, Fatoki JO, et al. (2022) Ellagic Acid and Hesperetin Ameliorates Lipopolysaccharide-Induced Multiple Organ Perturbation of Glycolytic and Mitochondrial Metabolizing Enzymes in Murine Model. FUOYE Journal of Pure and Applied Sciences. 7(1): 79-98.
View at Publisher | View at Google Scholar - Sanchez-Abarca LI, Tabernero A, Medina JM. (2001). Oligodendrocytes use lactate as a source of energy and as a pre-cursor of lipids. Glia, 36 (3): 321–329.
View at Publisher | View at Google Scholar - Bricker AL, Carey VJ, Wessels MR. (2005). Role of NADase in virulence in experimental invasive group A streptococcal infection. Infect Immun. 73(10):6562-6.
View at Publisher | View at Google Scholar - Heikal AA. (2010). Intracellular coenzymes as natural biomarkers for metabolic activities and mitochondrial anomalies. Biomarkers in Medicine. 4(2):241-263.
View at Publisher | View at Google Scholar - Xie N, Zhang L, Gao W, Huang C, Huber PE, et al. (2020). NAD+ metabolism: pathophysiologic mechanisms and therapeutic potential. Signal transduction and targeted therapy. 5(1):1-37.
View at Publisher | View at Google Scholar - McKenna MC. (2007). The glutamate–glutamine cycle is not stoichiometric: fates of glutamate in brain. Journal of Neuroscience Research. 85(15):3347–3358.
View at Publisher | View at Google Scholar - Gupta R, Gupta N. (2021). Tricarboxylic Acid Cycle. In: Fundamentals of Bacterial Physiology and Metabolism. Springer, Singapore. 2021.
View at Publisher | View at Google Scholar - Voet D, Voet JG, Pratt CW. (2015). Fundamentals of Biochemistry: Life at the Molecular Level (4th ed.). Hoboken, NJ: Wiley. 574–575.
View at Publisher | View at Google Scholar - Spanaki C, Zaganas I, Kleopa KA, Plaitakis A. (2010). HumanGLUD2Glutamate Dehydrogenase Is Expressed in Neural and Testicular Supporting Cells. Journal of Biological Chemistry, 285(22): 16748–16756.
View at Publisher | View at Google Scholar - Lyall Fiona. (2010).
View at Publisher | View at Google Scholar - Kracke F, Vassilev I, Krömer JO. (2015). Microbial electron transport and energy conservation - the foundation for optimizing bioelectrochemical systems. Frontiers in Microbiology. 6: 575.
View at Publisher | View at Google Scholar - Li LP, Liu XF, Ning YH, Liu GZ, Li CY. (2010). Effects of DEHP on rat testicular enzymes of energy metabolism and its role in oxidative damage. J. Ningxia Med. Uni. 32 (1): 74–77.
View at Publisher | View at Google Scholar - Lin DF, Hou XJ, Zhang WJ, Zhou JP, Li J. (2009). Effects of diphenyl methane diisocyanate exposure at gestation on energy metabolism in testis of adult male offspring mice. J. Environ. Health. 26 (4): 299–301.
View at Publisher | View at Google Scholar - Wu D, Li Y, Tang B, Fang F, Ding SM, Yang X, et al. (2007). Effects of butyl benzyl phthalate on enzymes related to energy metabolism in testis of mice. Asian J. Ecotoxicol. 2 (4): 440–444.
View at Publisher | View at Google Scholar - Park LC, Albers DS, Xu H, Lindsay JG, Beal MF, et al. (2001). Mitochondrial impairment in the cerebellum of the patients with progressive supranuclear palsy. Journal of neuroscience research. 66(5):1028-34.
View at Publisher | View at Google Scholar - Eric Schon, Giovanni Manfredi (2003) Neuronal degeneration and mitochondrial dysfunction. J Clin Invest. 111(3):303–312.
View at Publisher | View at Google Scholar
