

Review Article | DOI: https://doi.org/10.31579/2834-8532/019
Twenty Years of the Cell Thermoregulation Hypothesis
1 International Higher School of Medicine, Bishkek
2 National Center of Cardiology and Internal Medicine, Bishkek, Kyrgyzstan.
*Corresponding Author: Abyt Ibraimov, International Higher School of Medicine, Bishkek and National Center of Cardiology and Internal Medicine, Bishkek, Kyrgyzstan
Citation: Abyt Ibraimov (2023), Twenty Years of the Cell Thermoregulation Hypothesis, Clinical Genetic Research, 2(2); Doi:10.31579/2834-8532/019
Copyright: © 2023 Abyt Ibraimov,This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 17 April 2023 | Accepted: 24 April 2023 | Published: 19 April 2023
Keywords: cell thermoregulation; condensed chromatin; chromosomal heterochromatin regions; human body heat conductivity; obesity; alcoholism; drug addiction
Abstract
The role of temperature in biological life is obvious, because, with rare exceptions, life is possible only in conditions of positive temperature. All forms of life in one form or another have the ability to control body temperatures. Prokaryotes, in response to extremely high or low temperatures, produce special proteins to protect proteins from denaturation. Ectothermic animals are able to use external sources of thermal energy (the sun) to maintain a positive temperature in the body. Endothermic animals develop most of the heat mainly from their body’s metabolisms. The organ-based (organismal) thermoregulation system includes the hypothalamus in the brain, as well as the sweat glands, skin, and circulatory system. However, the question of the possibility of thermoregulation at the cellular level remains open. Based on the study of chromosomal heterochromatin regions (HRs) in higher eukaryotes, the hypothesis of the existence of thermoregulation at the cellular level was substantiated. The essence of the hypothesis of cell thermoregulation is to eliminate the temperature difference between the nucleus and the cytoplasm when, for one reason or another, the nucleus temperature becomes higher than in the cytoplasm. At the same time, higher eukaryotes use a dense layer of peripheral condensed chromatin, which is the highest form of HRs organization in an interphase cell, as a heat conductor to eliminate the temperature difference between the nucleus and the cytoplasm. Since individuals in the population differ in the quantitative and qualitative content of HRs, this circumstance may affect the speed of eliminating the temperature difference between the nucleus and the cytoplasm with all the ensuing consequences for the whole organism.
Introduction
All forms of life in one form or another have the ability to control body temperatures. Prokaryotes, in response to extremely high or low temperatures, produce special proteins (heat shock or cold shock proteins) to protect proteins from denaturation. Ectothermic animals are able to use external sources of thermal energy (the sun) to maintain a positive temperature in the body. Endothermic animals develop most of the heat mainly from their body’s metabolisms. The organ-based (organismal) thermoregulation system includes the hypothalamus in the brain, as well as the sweat glands, skin, and circulatory system. However, the question of the possibility of thermoregulation at the cellular level remains open.
Based on the analysis of the distribution of the amount of chromosomal heterochromatin region (HRs) in the genome of human populations living in various environmental conditions of Africa and Eurasia, in norm and pathology the hypothesis of the existence of thermoregulation at the cellular level was substantiated [1]. The essence of the hypothesis of cell thermoregulation is to eliminate the temperature difference between the nucleus and the cytoplasm when, for one reason or another, the nucleus temperature becomes higher than in the cytoplasm. The higher eukaryotes use a dense layer of peripheral condensed chromatin, which is the highest form of HRs organization in an interphase cell, as a heat conductor to eliminate the temperature difference between the nucleus and the cytoplasm. Since individuals in the population differ in the quantitative and qualitative content of HRs, this circumstance may affect the speed of eliminating the temperature difference between the nucleus and the cytoplasm with all the ensuing consequences for the whole organism.
It is obvious that the nucleus, unlike the cytoplasm, cannot transfer heat directly into the intercellular space, from where heat will be withdrawn by the circulating fluid (blood). Thus, the nucleus can transfer excess thermal energy only to the cytoplasm. At the same time, the nucleus has two possibilities: to increase its volume or to strengthen the heat-conducting ability of the nuclear envelope. Since the first option is limited, and the second is impossible due to the thickness of the cell membrane, it seems that higher eukaryotes took advantage of the possibility of a dense layer of peripheral condensed chromatin (CC) as a heat conductor to more effectively eliminate the temperature difference between the nucleus and the cytoplasm. CC, localized between the nucleus and cytoplasm, consists of chromosomal HRs, which are one of the forms of organization of non-coding, so-called "redundant" DNA in the genome of higher eukaryotes. The data confirming the existence of CT are discussed below on the example of a human.
Why is the possibility of thermoregulation at the level of cells not being paid due attention?
The lack of interest in the possibility of the existence of cell thermoregulation has quite objective reasons. Recognized internal sources of heat (cellular metabolism, muscle contraction, ion pump, in rare cases brown fat burning) in the body are localized in the cytoplasm. Organisms receive energy from the environment in the form of potential energy contained in the chemical bonds of molecules of fats, carbohydrates and proteins. Energy metabolism of cells occurs mainly in cytoplasm. Apparently, it is taken for granted that if the temperature in the cytoplasm rises above the optimal level for the body, then it will be freely dissipated into the intercellular space. The cell nucleus is usually not considered as one of the important internal sources of heat, despite the fact that very active biochemical processes take place there (reparation, recombination, rearrangement, modification, restriction, replication, transcription and other processes associated with DNA).
However, the question of whether thermoregulation can also exist at the level of individual cells remains open. Apparently, it is taken for granted that the level of intracellular temperature cannot rise to dangerous physiological values due to the small size of cells. If this happens, then the presence of fluid in the intercellular space and the circulation system will not allow the temperature in the cells to rise above the optimal temperature for the organism. Therefore, the level of thermal energy released in the process of cellular metabolism cannot be so important that in the process of evolution appeared some special mechanisms for thermoregulation at the level of individual cells. Obviously, for the same reasons, it is considered that the nucleus and cytoplasm use the same mechanisms in removing excess thermal energy. It is considered that diffusion and possibly convection are the primary means to passively remove the heat generated inside the cell [2]. However, there are facts and observations, seemingly unrelated, but indirectly indicating the possibility of thermoregulation at the cellular level. Below we will discuss some of them.
Why should due attention be paid to the possibility of thermoregulation at the level of individual cells?
Based on the study of distribution of chromosomal Q-heterochromatin regions (Q-HRs) in the genome of various human populations, in norm and at some forms of human pathology, the hypothesis was put forward about the possibility of thermoregulation at the level of cells [1].
Initially, the cell thermoregulation (CT) hypothesis was proposed as an attempt to explain
The following facts:
- consistent intra- and interpopulation differences in the quantitative content of chromosomal Q-HRs in the human genome;
- these differences proved to be related to features of the ecological environment of the place of permanent residence, and not to racial and ethnic composition of the population;
- the number of Q-HRs in the genome tend to decrease from southern geographical latitudes to northern ones, and from low-altitude to high-altitude ones;
- different age groups have different number of Q-HRs, the greatest number of Q-HRs is characteristic of neonates, while the lowest – of elderly subjects;
- individuals capable of successfully adapting themselves to the extreme high-altitude climate (e.g., mountaineers) and of the Far North (e.g., oil industry workers of polar Eastern Siberia) are characterized by extremely low amounts of Q-HRs in their genome;
- individuals with a lower amount of Q-HR in their genome proved to be prone to alcoholism and obesity, while those with a greater amount of Q-HR – to drug addiction [1,3-21].
However, over time it became conceivable that some other facts and observations that still have no explanation could be partially explained within the framework of the CT hypothesis, in particular:
a) the existence of "excess" DNA in the genome of higher eukaryotes and why they consist of non-coding, repeating sequences of nucleotides;
b) the organization of a part of the "excess" DNA into chromosomal HRs, which make up from 10% to 60% of the DNA of the genome in higher eukaryotes;
c) formation of chromosomal HRs a dense layer of condensed chromatin (CC) around the nucleus;
d) the role that chromosomal HRs play is still essentially unknown. This is also reflected in the variety of hypotheses. These range from the idea that heterochromatin has no function to the assumption that it has an important role in development and evolution. However, none of them backed up by solid evidence when it comes to demonstrating their phenotypic manifestation;
e) the origin of homoeothermic animals;
f) the emergence of nucleolus and chromocenters in interphase nucleus;
g) the role chromosomal bands (C, G, Q and R) in development and evolution. It has been shown that G-, Q- and R-bands are absent in plants and are always present in the chromosomes of higher vertebrates. In the case of invertebrates, fish and amphibians, it is difficult to identify these bands. Some chromosomal bands in insects are equivalent to C-segments, and G-, Q- and R-bands appear to be absent. With the help of differential staining of chromosomes in plants, it is possible to identify mainly C-bands. However, chromosomal bands are best detected in the karyotype of higher primates, especially in humans;
h) the biological meaning of the presence only in the karyotype of three higher primates (Homo sapiens, Pan troglodytes and Gorilla gorilla) a new type of constitutive heterochromatin - Q-heterochromatin;
i) the existence of a broad quantitative variability of chromosomal Q-HRs only in the genome of human populations;
j) the origin in the karyotype of modern man of 46 instead of 48 chromosomes;
k) high physiological plasticity of a man, which allowed him to master the whole land of the Earth.
l) a man is able to run continuously for such long distances that no homoeothermic mammalian can do.
What is the cell thermoregulation?
To better understanding of cell thermoregulation (CT) hypothesis, it is necessary to briefly recall some facts about chromosomal HRs in the genome of higher eukaryotes. To-date two types of constitutive heterochromatin are recognized: C- and Q-heterochromatin [22,23,24]. There are significant differences between them: C-heterochromatin is found in the chromosomes of all the higher eukaryotes, while Q-heterochromatin - only in man, the chimpanzee and gorilla [25-27]. C-HRs are known to be invariably present in all the chromosomes of man, varying mainly in size and location (inversion). Q-heterochromatin regions (Q-HRs) may be completely absent on any of the Q-polymorphic chromosomes without any noticeable phenotypic consequences for the carrier, while the complete absence of C-HRs even on one chromosome is an extremely rare phenomenon [24].
Only seven pairs of autosomes (3, 4, 13-15, 21 and 22) and the Y chromosome can have Q-HRs in the human karyotype. The main feature of the phenomenon of wide variability of Q-HRs is that individuals in populations differ in the number, localization, size and intensity of fluorescence. It should be emphasized that there is no individual in human populations who would have Q-HRs simultaneously on all 25 potentially polymorphic loci of 46 chromosomes in his karyotype. Usually, the number of Q-variants in the human karyotype in populations ranges from 0 to 10 [14].
Here it is necessary to emphasize once again that: a) the broad quantitative variability of chromosomal Q-HRs is present only in the genome of human populations, despite the fact that they occur in the genome of two more higher primates; b) human populations do not differ from each other in the quantitative content of chromosomal C-HRs [10,30].
Now about what we mean by the CT. CT refers to the process of equalizing the temperature difference between the cytoplasm and the nucleus. The inner surface of the nuclear membrane is occupied by heterochromatin in the form of a dense layer of condensed chromatin (CC) and lamina (Figure. 1).
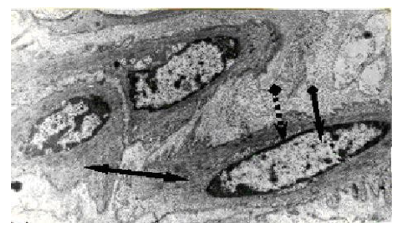
Figure 1: Electron micrograph of smooth muscle cells of a bull (x 8 000). Condensed (heterochromatin) and decondensed chromatin (euchromatin); ∙∙∙∙> peripheric layer of condensed chromatin; → euchromatin; and ↔ cytoplasm can be seen in the nuclei [28].
We believe that the structural basis of CT is the peripheral layer of CC. The transfer of thermal energy between the cytoplasm and the nucleus is carried out through this dense layer of peripheral CC, located inside the nuclear envelope [1].
Temperature homeostasis in the nucleus is necessary, first of all, for the optimal flow of complex molecular processes associated with the metabolism of DNA and RNA, which are important for the synthesis of normal proteins. Therefore, the hypothesis postulates that the cell strives to maintain a temperature within very narrow limits. As is known, chromosomes have internal (reparation, recombination, rearrangement, modification, restriction) and external (replication, transcription, packaging, organized movement) activities, which are accompanied, among other things, by the release of thermal energy. If, for some reason, the temperature in the nucleus begins to exceed that in the cytoplasm, there is a need for dissipation of excess heat outside the nucleus.
We believe that unlike the cytoplasm the nucleus is in the most difficult position in maintaining temperature homeostasis. Indeed, in order to get rid of excess heat, the nucleus has two choices: increase in size and/or increase the density of the nuclear envelope to speed up the transfer of heat to cytoplasm. The first way to dissipate excess heat from the nucleus is difficult due to the fact that the cytoskeleton and intracellular organelles do not allow this. The second choice is possible if the nucleus can increase the density of the CC in order to increase its heat conductivity.
It is obvious that the thickness and density of the CC depends on the number of chromosomal C-HRs in the karyotype. However, in the genome of three higher primates, unlike other animals, in addition to C-HRs, there are also Q-HRs, which, as we believe, capable to further increase the compactization of CC. In other words, CC in three higher primates (which contains chromosomal C- and Q-HRs), being the most densely packed material, should have the highest heat conductivity in the interphase cell with all the ensuing consequences [1,29,31-41].
But this does not mean that CT is exclusively a product of chromosomal HRs. In the chromosomes of endothermic animals in addition to constitutive HRs there are other tightly packed regular areas, like G+ and Q+ bands. In the composition of the chromosomal G+ and Q+ bands there are heterochromatin materials (intercalary heterochromatin).
We also believe that in the dissipation of heat surplus from a nucleus a nucleolus is actively involved. Thus, some of the chromosomes have nucleolar organizer regions (NORs), which contain ribosomal cistrons. In humans, NORs are localized on five pairs of acrocentric chromosomes (13-15, 21 and 22). As it is known, NORs, together with chromosomal HRs form the nucleolus bodies. Since HRs are in the body and around the nucleolus, there is nothing surprising in the assumption that they can promote removal of excess heat from the nucleolus and further transfer to CC around the nucleus [42,43].
It has long been established that in the nucleus chromosomal HRs coalescent into a chromocenters. However, their biological role is still unclear. Existing hypotheses link the possible biological role of chromocenters with the behavior of HRs in the cell cycle. We believe that the nuclear areas where excess heat is generated form temporary structures in the form of chromocenters and nucleoli, connecting them with a peripheral layer of СС. The essence of the proposed hypothesis is the assumption that the chromocenters, along with the nucleolus and the dense layer of peripheral СС participate in СТ. Namely, chromocenters are involved in the dissipation of excess heat from the "hot" areas of the interphase nucleus [42-44].
We also believe that the genetic inertness of chromosomal HRs may not be the only reason for their localization on the nuclear periphery. It seems highly probable that the dense packing of HRs is caused by two reasons: a) the need for CT to remove excess heat from the nucleus; and b) the risk of damage the structure of the nuclear membrane from exposure to high temperature. The first reason, apparently, does not need any additional arguments, since the CC located on the nuclear periphery is the densest and, accordingly, the most highly heat-conducting structure in the interphase cell. The second reason is related to the features of the cell membrane. As it is known, cell membranes are very sensitive to temperature fluctuations: they become too solid at low temperatures and too liquid at high temperatures to perform their functions normally. Of course, to remove excess heat from the nucleus, a layer of cell membrane would be ideal due to their thickness. However, the high vulnerability of cell membranes to temperature fluctuations apparently “forced” Nature to use a dense CC layer of HRs, lamina, nucleolus, chromocenters and cytoskeleton to protect their functional integrity. The lamina is located between the layer of CC and the nuclear envelope. For what reason? It is generally accepted that the lamina exists below the inner nuclear membrane to give strength and shape to the nucleus. But, in principle, the lamina could be located outside the nuclear envelope, like a cell wall in plant cells or in prokaryotes, if the task of the lamina is limited only to strengthening and shaping the nucleus. We believe that there is a biological meaning in the localization of the lamina under the inner nuclear membrane - to protect the nuclear envelope from the harmful effects of heat emanating from the biochemically highly active nucleus.
Despite the fact that the nucleoli, chromocenters and B-chromosomes were discovered more than a hundred years ago, their biological role is still not completely clear. We believe that in order to protect the nuclear envelope from the deleterious effects of heat emanating from the nucleus, nucleoli and chromocenters are formed, and in some cases, Nature uses the services of B-chromosomes. The essence of our hypothesis lies in the assumption that the nucleoli, chromocenters and B-chromosomes, consisting of constitutive heterochromatin, are involved: a) in the localization of “hot” areas in the nucleus; b) in the direction of the “flow” of heat to very limited areas of the nuclear membrane, in order to protect the functional integrity of the nuclear membrane. In other words, if heat is removed evenly around the circumference of the nucleus, then there may be a danger of damage to the entire nuclear envelope [28,35,42-44].
The observations that human autosomes 1, 9 and 16, which do not contain ribosomal cistrons but have largest blocks of HRs and are in contact with the nucleolus [46], still have no explanation. The predominant spatial proximity of the sex chromosomes to the nucleoli is in the same situation [47]. As it seems to us, a rational explanation of these facts is the assumption of their participation in the "localization" of excess heat in the nucleolus, and then contacting with CC to dissipate it into the cytoplasm [42,44].
There is no known mechanism for a cell to dissipate excessive heat. It is considered that diffusion and possibly convection are the primary means to remove the heat generated inside the cell. However, this explanation is strongly objected to. According to Albrecht-Buehler [45] ‘Inside the cell the molecules are mostly associated with polymeric structures (cytoskeletal polymers or membranes) and thus exist in very heterogeneous, solid state environments that alter their behavior dramatically compared to free molecules in test tubes’. In other words, the molecules in the cell do not float freely, as with a water solution. Therefore, diffusion and convection cannot be the primary mechanisms to remove the excess thermal energy generated inside the cell. Consequently, it is necessary to look for other additional mechanisms for dissipation surplus heat from the cell, and especially from its largest organelle - the nucleus.
Experimental verification of the cell thermoregulation hypothesis.
The CT hypothesis has not yet been directly experimentally confirmed. Of course, it would be ideal if someone were able to show the following in vivo: the rate of heat transfer from the nucleus to the cytoplasm depends on the number of chromosomal HRs. No one has done such an experiment yet. However, the ultimate goal of our study is to experimentally test the CT hypothesis, which, as we believe, is the main biological effect of chromosomal HRs [1].
The idea of CT refers to the process of equalizing the temperature difference between the cytoplasm and the nucleus. Ultimately, we are talking about the heat conductivity of the entire cellular part of the body. The simplest idea of the body heat conductivity (BHC) can be represented on the basis of the known laws of physics. It follows from the second law of thermodynamics that heat transfers from a hot body to a cold one until the temperature difference disappears. It is also obvious that the same substance, depending on its density, should have different heat conductivity. The higher the density, the higher the heat conductivity, that is, the higher the rate of transfer of heat. CC is the densest area in the interphase cell, therefore, all other things being equal, it should have the highest heat conductivity.
Heat conductivity (HC) due to the transfer of energy is one of the three transfer phenomena existing in nature (heat conductivity, diffusion and internal friction or viscosity). All substances have HC: gases, liquids and solids. Unlike gases and liquids, convection is not possible in solids, therefore, the thermal transfer is carried out only by heat conductivity.
In thermal physics, the measurement of HC of solids (for example: metal) is carried out by determining the coefficient of thermal conductivity by the calorimetric method. The thermal transfer takes place through a metal rod, the ends of which are placed in a calorimeter with water taken at temperatures Т1 and T2 (T1 > T2). It is necessary to experimentally determine the amount of heat transferred and the time to measure the coefficient of HC of a given metal rod.
It is obvious that the direct transfer of the HC measurement method used in thermal physics is not acceptable for the human body, both for technical and ethical reasons. However, we tried to approach the solution of this problem indirectly, by assessing the HC of only a part of the human body. To do this, we had to modify the generally accepted technique of physicists so that it was acceptable to humans.
It is obvious that the living body has some initial heat conductivity. Nevertheless, as an important physical characteristic of a living organism, it has not yet attracted the attention of scientists. We failed to find in the literature not only a special method, but even no attempts to evaluate the BHC of living organisms in vivo. This is not surprising, because it turned out that even experts in the physiology of thermoregulation never raised this issue [48,49]. However, all this can be understood, because the animal body is not a homogeneous physical mass. Since direct in vivo measurement of the temperature of different parts of cells is still not possible, then it was decided to somehow indirectly estimate the heat conductivity of the whole body.
Perhaps the most difficult methodological problem in determining the human BHC was the creation of a temperature gradient between the human body and the source of thermal load individually for each person. It is known that people differ from each other in the temperature of the palm, and we decided to take advantage of this circumstance. Our empirical experience has shown that in order to create a temperature gradient between the human body and the source of thermal load, an acceptable value by which it is possible to painlessly increase the temperature of “hot” water in a water bath is ~9 °C. Of course, this number does not hide any fundamental physical characteristic of the human body. But it is also not desirable to increase the temperature of “hot” water by more than 9 °C. In this case, we could encounter, in addition to unpleasant sensations, the denaturation of proteins in cells in individuals whose temperature of the palm is close to human core temperature.
It has been found that the temperature of the palm itself, and especially when taken into account in close connection with the temperature of the forehead surface can give very informative results about the human BHC [41]. It is by studying the distribution of the temperature difference between palm and forehead surface we managed to identify the existence of significant differences in gender and racial and ethnic origin at the population level [28,41].
The main difficulty in searching for a possible relationship between the number of chromosomal HRs and the human BHC was the complete absence of any methodological approach in this regard. If the determination of the number of HRs in the human karyotype is already a well-established methodological procedure, then this cannot be said when it comes to measuring the human BHC due to the complete lack of any experience in this matter. Therefore, it is too early to say that we can measure the human BHC with the same accuracy as physicists do when measuring the thermal conductivity of nonliving bodies, such as metals. At best, we can only evaluate the human BHC as high, medium, or low, and apparently, it is still far from an accurate measurement in numbers [28,32,41].
Over the years, we have tested many methods of evaluating of the human BHC. By trial and error, we managed to significantly improve and simplify the technique of measurement of human BHC with the involvement of some technical tools and approaches. In particular, we began to use a water bath with a thermostatic device (water bath WTB6 from MEMMERT GmbH + Co., Germany) and pyrometers for non-contact thermometry of the surface of the human body.
The measurement of the human BHC is carried out indoors at room temperature (20 °С-24 °С). At first, the temperatures of the surface of the forehead and the palm of the hands were measured in the studied individuals. To prepare “hot” water for a given individual the number 9 added to the temperatures of the surface of the left palm. For example, if the temperature of the palm of the individual's left hand is 31.0 °C, then the temperature of the “hot” water should be 40.0 °C. The studied individual sat on a chair with a back, the torso was straightened, the head was raised, the arms hung naturally at the sides, and the muscles were relaxed. Then the individual slowly immersed the left hand up to the wrist in a water bath filled with “hot” water. Every minute, the temperature of the surface of the right palm of the studied individual was measured with a pyrometer to the nearest tenth of degrees Celsius (t °С). The experiment continues (thermal load of the human body through the left hand) until the temperature increase in the surface of the right palm reaches its maximum value (“temperature peak”) and begins to decrease (for details see [28,41]). How do we interpret these data? We believe that the time of the onset of the temperature peak on the surface of the right palm reflects the rate of heat transfer in the human. In other words, we believe that a human with high BHC is able to: a) conduct heat through his body faster than other individuals in the population; b) equalize the temperature difference in different parts of the body faster; and c) dissipate excess heat outside of his body faster in order to maintain the optimal temperature level in the body. If the temperature peak on the surface of the right palm occurs within the first 5 minutes from the start of the thermal load, then such an individual is considered as a human with a high BHC, from 6-10 minutes, as an average one, from 11 minutes and above, as a low BHC [41].
In the process of testing the method, it also turned out that there is a statistically significant relationship between the level of human BHC and the magnitude of the temperature difference between distant parts of his body: e.g., forehead and palm or armpit and palm surface temperature. We believe that the smaller temperature difference between different parts of the human body reflects the high heat-conducting capacity of this organism in the sense that: a) such an organism equalizes the temperature differences between different parts of the body more efficiently; b) dissipates excess heat energy outside of the body faster. As it turned out, that the temperature of the surface of the palm at rest also reflects the BHC level; individuals with a high palm temperature have a high BHC, and vice versa [41].
Table 1 shows the relationship between the number of chromosomal Q-HRs and the rate of reaction of the body to the controlled thermal load, which was determined by the time (in minutes) to the occurrence of peak temperature on the surface of the right palm.
| Number of Q-HRs | 1 to 5 minutes (n = 34) I | 6 to 10 minutes (n = 75) II | 11 minutes & > (n = 27) III | Total (n = 136) |
| 2 | 14 | 5 | 19 | |
| 3 | 2 | 12 | 9 | 23 |
| 4 | 5 | 29 | 8 | 42 |
| 5 | 14 | 7 | 4 | 25 |
| 6 | 8 | 3 | 1 | 12 |
| 7 | 3 | 7 | 10 | |
| 8 | 2 | 3 | 5 | |
| Total Q-HRs | 181 | 306 | 95 | 582 |
| Mean number of Q-HRs |
5.32 ± 0.206 |
4.08 ± 0.189 |
3.51 ± 0.209 | |
Statistics t I, II = 3.975; df = 107; P = <0> t II, III = 1.656; df = 100; P = 0.101; t I, III = 6.083; df = 59; P = <0> | ||||
Table 1: The distribution and mean numbers of chromosomal Q-HRs and time to occurrence of temperature peak on the surface of the right palm.
These differences are statistically significant.
As can be seen in this Table, there is a statistically significant relationship between the number of chromosomal Q-HRs in the human genome and the reaction of the body to the controlled thermal load. In individuals whose genome contains more chromosomal Q-HRs than the population average, the
peak temperature occurs in the first five minutes of the thermal load, and vice versa. The relationship between the quantity of chromosomal Q-HRs and the temperature difference between the surface of the right palm and the forehead at rest is shown in Table 2.
Number |
of |
0.1 °C to |
to |
2.1 °C to |
Total | ||
| Q-HRs | 1.0 °C | 2.0 °C | 3.0 °C | (n = 136) | |||
| (n = 32) | (n = 71) | (n = 33) | |||||
| I | II | III | |||||
| 2 | 10 | 9 | 19 | ||||
| 3 | 12 | 11 | 23 | ||||
| 4 | 9 | 26 | 7 | 42 | |||
| 5 | 9 | 14 | 2 | 25 | |||
| 6 | 7 | 4 | 1 | 12 | |||
| 7 | 5 | 3 | 2 | 10 | |||
| 8 | 2 | 2 | 1 | 5 | |||
| Total Q-HRs | 174 | 291 | 117 | 582 | |||
| Mean number of Q-HRs |
5.44 ± 0.220 |
4.10 ± 0.168 |
3.54 ± 0.275 | ||||
| Statistics | t I, II = 4.607; df= 101; P = <0> t II, III = 1.786;df = 102; P = 0.077; t I, III = 5.349; df = 63; P = <0> | ||||||
Table 2: The distribution and mean numbers of chromosomal Q-HRs and the temperature difference between the surface of the right palm and the forehead.
These differences are statistically significant.
As can be seen in Table 2 the more chromosomal Q-HRs, the smaller the temperature difference between the forehead surface and the right palm, and vice versa. Table 3 shows a different pattern: the greater the number of chromosomal Q-HRs, the higher the temperature of the surface of the right palm at rest, and vice versa.
Number of Q-HRs | Below 35.0 °С (n = 36) I | 35.1 °С to 36.0 °С (n= 74) II | 36.1 °С and over (n = 26) III | Total (n = 136) |
| 2 | 5 | 14 | 19 | |
| 3 | 6 | 17 | 23 | |
| 4 | 16 | 23 | 3 | 42 |
| 5 | 7 | 7 | 11 | 25 |
| 6 | 2 | 5 | 5 | 12 |
| 7 | 6 | 4 | 10 | |
| 8 | 2 | 3 | 5 | |
| Total Q- HRs | 139 | 294 | 149 | 582 |
Mean number of Q-HRs |
3.86±0.179 |
3.97±0.185 |
5.73±0.239 | |
Statistics t I,II = 0.380; df= 108; P = 0.704; t II,III = 5.111; df = 98; P = <0> t I,III = 6.395; df = 60; P= <0> | ||||
Table 3. The distribution and mean number of chromosomal Q-HRs and the temperature of the surfaceof the right palm.
These differences are statistically significant.
How do we interpret these data? We believe that the time to occurrence of peak temperature (T) on the right palm reflects the rate of heat conductivity, while the value of T of the right palm’s surface at that moment seems to reflect the quantity of thermal energy in the individual’s body. If the peak T on the surface of the palm occurs in the first five minutes after the thermal load is applied, then such an individual is considered to have a high BHC, and vice versa.
A statistically significant relationship between the number of chromosomal Q-HRs and the T difference between the forehead surface and the right palm at rest may also characterize the human BHC — the smaller the T difference, the higher the BHC, and vice versa. We believe that a smaller T difference between the forehead surface and the palm reflects the high heat conductivity of the body, so that such an organism equalizes the T difference between the different parts of the body more effectively, thereby successfully avoiding overheating in high temperature conditions. The temperature of the right palm at rest, presumably, also reflects the level of BHC: individuals with a high T at the palm may have higher BHC, and vice versa.
What does the CT hypothesis offer to explain?
From our point of view, the CT hypothesis can explain the above facts, provided that: a) there really is a problem with the removal of excess metabolic heat from the cell nucleus; b) the peripheral CC consisting of chromosomal HRs is a best heat conductor between the nucleus and the cytoplasm; c) the packing density of the CC depends on the number of Q-HRs; d) individuals in the population really differ from each other in their BHC level and it depends on the number of chromosomal Q-HRs in their genome; e) the mechanisms of human physiological thermoregulation are the same, but they are implemented under different physical conditions due to the fact that the bodies of individuals in the population differ in their level of heat conductivity with all the consequences that follow from this.
Let's start by analyzing: a) the consistent intra- and inter-population differences in the quantitative content of chromosomal Q-HRs in the genome of human populations living in different ecological environments; b) the tendency to decrease the number of Q-HRs in the human genome from southern geographical latitudes to northern and from low to high altitude ones; c) the number of Q-HRs in people who are able to successfully adapt to the extreme high-altitude climate and the Far North, in individuals with alcoholism, obesity and drug addiction. We want to assert that the individuals with a high BHC equalize better and faster temperature differences between different parts of the body, and vice versa. If that is the case, then, for example, the well-known resistance of the natives of the southern latitudes to high ambient temperatures would find a rational explanation. Namely, southerners, due to the high heat conductivity of their bodies, more effectively equalize temperature differences in different parts of the body and quickly dissipate excess heat into the environment. In this case, the natives of the Far North and high altitudes can better and longer retain metabolic heat in the organism due to the low heat conductivity of their bodies, with all the ensuing consequences for their adaptation (for more details, see [10-15,28,50]).
The facts of the low number of chromosomal Q-HRs in the genome of older people compared to newborns and children [5,51,52] can be explained within the framework of CT hypothesis. We believe that the decrease in the number of Q-HRs in older age groups is associated with the negative selection of children in the first days, months and years of life, in which the levels of BHC are high for a given ecological environment. It is known that the small weight and relatively large surface of the child’s body lead to a discrepancy between heat production and heat dissipation. The younger the child, the greater the body area per unit of body weight and, consequently, the greater its heat loss. At the same time, 0.069 m2 of body surface falls on 1 kg of a child's weight, while in an adult - 0.025 m2, i.e., the cooling surface of a child is more than twice that of an adult per 1 kg of weight. Moreover, in a newborn, 2/3 of the entire circulating blood is in the skin. This is also largely due to the huge surface of the child's skin in relation to body weight. Since it is known that in the first years of life, children die in the absence of timely and effective medical care, mainly from respiratory diseases, as well as infections due to hyperthermia. The same probably occurs in subsequent years of life, since it is known that during ontogenesis the number of chromosomal Q-HRs does not change.
Let us briefly consider the situation with alimentary obesity. We found that individuals suffering from alimentary obesity have a significantly lower BHC compared to the control sample [28,33.35]. The main question remains open – why are some individuals thin and others fats even in a relatively homogeneous environment? It is believed that the answer to this question will be obtained by identifying the genes responsible for obesity in humans. With respect to obesity, we believe the following: in patients with alimentary obesity with a low number of Q-HRs, under favorable conditions and material standard of living, part of the calories in a low heat-conducting body accumulates in the form of body fat due to poor dissipation of excess metabolic heat from the body.
We would answer the notorious question: “Why are some individuals thin and others very fat?” instead of existing points of view that obesity is the result of a lack of internal discipline in food ingestion or the presence of genes responsible for obesity, that obesity is not just a personal failure or the result of a malfunction of any structural genes. We believe that, apparently, there is a wide diversity in the functioning of energy and temperature homeostasis and this diversity is associated with the BHC level of the individual. In individuals with low BHC, even when consuming the same amount of food as people with normal weight, under comfortable living conditions, more fat will be deposited due to a smaller number of chromosomal Q-HRs, which, as we believe, are involved in human CT thermoregulation as part of CC in the nucleus [1,31-33].
Alcoholism and drug addiction are exclusively human pathologies. Our data show that the number of chromosomal Q-HRs in the genome seems to have something to do with the pathogenesis of alcoholism and drug addiction. As we have shown, in the genome of individuals who abuse strong alcoholic beverages, the number of Q-HRs is extremely small, while at the same time, people with drug addiction, on the contrary, have a lot of Q-HRs [34,35].
The possible role of BHC in the development of alcoholism appears to us as follows; the frequency of consuming strong alcoholic beverages tends to increase with increasing latitude (from low to high) and altitude. At the same time, the number of Q-HRs in the genome in a population tends to decrease as the geographical latitude increases and the height of the place of permanent residence above sea level increases [1,9,14,28].
Let us consider the simplest example. Often life and the harsh climate in the Far North or the high-altitudes predispose, in a certain sense, to the intake of strong alcoholic beverages just to obtain a feeling of thermal comfort. But at the same time, as we believe, the same dose of alcohol for people with different BHC can lead to different consequences. So, a feeling of thermal comfort in individuals with low BHC occurs after in taking a relatively large amount of alcohol in one feast due to delayed “warming up” and equalization of the temperature difference in different parts of the body, which eventually leads to more severe intoxication with a hangover syndrome than in individuals with normal or high BHC. In other words, the lower the BHC of an individual, the stronger alcohol is required due to the slow warming of the whole body necessary for the onset of a feeling of thermal comfort in the whole body.
Addiction to drugs in drug addicts, that is, individuals with high BHC also occurs due to the desire to get a sense of thermal comfort., but this “pleasure” specifically comes from the “narcotic cooling” of the body, with the resulting emotional or other sufferings. We believe that the psycho-emotional effects of alcohol and drugs on the body are determined by the depth of violation of temperature homeostasis at the cellular level, but manifested in diametrically opposite directions, that is, ethanol causes alcohol intoxication, raising body temperature (the oxidation of 1 g of ethanol produces 7 kcal), and drugs, on the contrary, lower it, causing a state of narcotic stupor.
The natural human desire in a hot climate to enjoy the “deep coolness” or thermal comfort in the North or the highlands would be a completely justified desire if they were not satisfied with narcotic stupor or alcohol intoxication. However, the notorious propensity of southerners to take drugs, and northerners or highlanders to strong alcoholic beverages, could be partly explained by the different content of Q-HRs in their genome [6,11-13,16-18] and, accordingly, associate them with the human BHC [28,33,34].
Regarding the origin of homoeothermic animals, let us recall once again that maintaining the relative constancy of the internal temperature is a necessary condition for normal life. Some living beings maintain temperature homeostasis in the body due to external sources of energy (poikilothermy), others due to the energy of food consumption (homeothermy). However, it is unknown the origin of homoeothermic organisms. Despite the fundamental similarity of the mechanisms of the central organ-based physiological thermoregulation, even among the higher vertebrates exists poikilothermy and homoeothermic animals. We believe that homeothermy is not the result of the evolution of physiological mechanisms of thermoregulation. Homeothermy is the result of the evolution of high repetitive DNAs in the genome, some of which formed the chromosomal HRs. HRs constitutes the material basis of CT, which is responsible for the removal of excess thermal energy from the nucleus into the cytoplasm. Homoeothermic organisms, unlike poikilotherms have chromosomal G-, Q- and R-bands and capable of faster and more efficient leveling of temperature difference between the nucleus and the cytoplasm with all the ensuing consequences [28,39 40,43].
It has been shown that chromosomal G-, Q- and R-bands are absent in plants and are always present in the karyotypes of higher vertebrates (reptiles, birds and mammals). In the case of invertebrates, fish and amphibians, it is difficult to identify these bands. Chromosomal bands (CBs) are dense regions of the chromosomes. Based on study of CBs the hypothesis about their participation in the CT has been presented. The essence of hypothesis is that the CBs as part of peripheral layer of CC serves as heat conductor for a more efficient elimination of the temperature difference between the nucleus and cytoplasm [43,44].
We do not know the cause of the origin only in the karyotype at the ancestors of three higher primates (Homo sapiens, Pan troglodytes and Gorilla gorilla) a new type of constitutional heterochromatin - Q-heterochromatin. In addition, the origin of 46 instead of 48 chromosomes in the karyotype of modern man, characteristic of higher primates, is still not the subject of special research.
The cause and effect of such chromosome rearrangements is unknown. A hypothesis has been proposed that natural selection caused merger of two pairs of autosomes into one chromosome. In the changed climate of the East Africa individuals with a smaller number of chromosomal Q-HRs were the most adapted. Two pairs of acrocentrics in the genome the ancestor of modern man, which merged into a single chromosome, apparently, carried on their short arms of Q-HRs with a very high frequency, preventing the birth of individuals with a low number Q-heterochromatin. With the merger of these two pairs of acrocentrics into one, the number of autosomes bearing the Q-HRs reduced from nine to seven pairs, as in the modern human. Such chromosome rearrangement resulted in two important consequences: а) chromosomal Q-HRs distributed into seven Q-polymorphic autosomes, so that it was possible to give birth to the individuals with different, including the low number of Q-heterochromatin; b) in the population individuals with low number of Q-HRs appeared, able to adapt to new, harsher climatic conditions. With the lapse of time, these individuals formed a new population in the new territory, where individuals with a number of Q-HRs like the modern man, and with the number of 46 chromosomes in the genome began to dominate. Thus, the cause of the origin of the 46 chromosome karyotype from an ancestral 48 chromosome line was natural selection, and an effect was adaptation, i.e. individuals with different, including the low number of Q-HRs, got the advantage to open up and to colonize new ecological zones of the East Africa (for details see [36,40]).
Speaking about the high physiological plasticity of a human, which allowed him to adapt to climatic conditions other than East Africa and to master the whole land of the Earth, we mean the following. In fact, there is nothing special behind the figure 46. There are many animals and plants with this number of chromosomes. Here, in our opinion, the localization, composition, and especially wide quantitative variability of chromosomal HRs in the human karyotype are important. The uniqueness of the human karyotype is as follows: 1) unlike other animals only humans and two other higher primates have both types of constitutive heterochromatin - C- and Q-heterochromatin; 2) among higher primates, the largest number of chromosomal C-HRs is in the human karyotype, and they are localized on its three autosomes (1, 9 and 16) and on the Y chromosome. A human owes the highest heat conductivity of his body to this very circumstance; 3) in contrast to chimpanzees and gorillas, the number of Q-HRs in individuals in a population varies and ranges from 0 to 10; 4) such a wide quantitative variability is associated with an uneven distribution of the number chromosomal Q-HRs on seven Q-polymorphic autosomes; 5) the phenotypic manifestation of such hereditary variability is the differences between individuals in a population from each other in terms of BHC levels with all the ensuing consequences for the organism.
As is known, a man is able to run continuously for such long distances that no homoeothermic mammalian can do (e.g., corral hunting, when a primitive hunter in the conditions of African heat can drive an animal to death). In short, from our point of view this curious circumstance is due to the uniqueness of the karyotype of modern man. Namely, only man has the highest BHC among mammals, since in his karyotype, in addition to Q-HRs, there are autosomes (1, 9 and 16) that carry the largest blocks of C-HRs among higher primates.
Within the framework of the CT hypothesis, other well-known facts could be explained. For example, more and more countries located in low geographical latitudes began to take part in international sports competitions. It is noteworthy that athletes living in these regions achieve great success in sports that require (in addition to other factors) efficient heat dissipation from the body (e.g. football, professional boxing and marathon running). In contrast, athletes from higher geographic latitudes predominate in water sports, winter sports, and alpinism [16,17,29,38]. It has been established that natives of low geographical latitudes have more chromosomal Q-HRs in their genome [6,11-14,18,20]. Because southerners' bodies have relatively high BHC level [32], it is not surprising that they are successful in sports requiring efficient heat dissipation. Indeed, an athlete with a high BHC level cannot make much progress in water and winter sports due to the fact that his body will cool too quickly. However, this same athlete is likely to be more successful in sports that require efficient heat removal from the body. In the same way, it would be possible to explain why men tolerate heat stress better than women, and the latter are more resistant to cold than men, since men have a total number of chromosomal HRs on average twice as much as women [28].
Conclusion
The biological role of “excess DNA” in eukaryotes, which consists of short repeating sequences of nucleotides and does not code for proteins and enzymes remains unclear. The part of this DNA in the interphase cell is complexed with proteins into highly compacted structures, referred to as CC. The purpose of the CT hypothesis is to provide data, that it is possible that in higher eukaryotes the biological role of a part of non-coding DNA, which form a dense layer of CC around the cell nucleus, is their participation in maintaining intracellular temperature homeostasis. Although the issue of thermoregulation at the cellular level is not yet on the agenda of scientists, nevertheless, an attempt has been made to collect information indicating the possibility of the existence of such a phenomenon. Since direct in vivo measurements of temperature changes in different parts of the cell are not yet technically possible, indirect methods by studying of parts of the human body and the number of heterochromatic regions of chromosomes in his genome are proposed to evaluate the possibility of thermoregulation at the cellular level. It is assumed that the phenotypic manifestation of CT of the human organism is the level of heat conductivity of his body, which role is to be assessed in normal and pathological conditions. Since the number of chromosomal HRs in the genome does not change in ontogenesis, then the BHC level may turn out to be the same constitutional feature as skin color, eye, body types, height and other physical human features.
Acknowledgements
I express my sincere gratitude to the colleagues who worked with me over the years – E.I. Aksenrod, G.U. Kurmanova, G.O. Karagulova, A.A. Akanov, T.S. Meimanaliev, M.T. Sultanmuratov, I.K. Moldotashev and A.K. Kazakova.
I apologize to those authors, whose works were not cited, or were cited only through reviews, owing to space limitations.
Conflicts of interest
none.
Funding
Funding has not been received for the study.
Statement of Consent/Ethical approval
Not required.
References
- Ibraimov AI. 2003. Condensed chromatin and cell thermoregulation. Complexus. 1: 164-170.
View at Publisher | View at Google Scholar - Hochachka PW. 2003. Intracellular Convection, Homeostasis and Metabolic Regulation. J ExpBiol, 206: 2001-2009.
View at Publisher | View at Google Scholar - Geraedts JPМ, Pearson P L. 1974. Fluorescent chromosome polymorphism: frequencies and segregation in а Dutch population. Clin Genet. 6: 247-257.
View at Publisher | View at Google Scholar - Müller НJ, Klinger НP, Glasser М. 1975. Chromosome polymorphism in а human newborn population. II. Potentials of polymorphic chromosome variants for characterizing the idiograms of an individual. Cytogenet Cell Genet. 15: 239-255.
View at Publisher | View at Google Scholar - Buckton КЕ, О’Riordan МL, Jacobs PА, et al. 1976. С- and Q-band polymorphisms in the chromosomes of three human populations. Аnn. Нum. Genet. 40: 90-112.
View at Publisher | View at Google Scholar - Lubs HА, Patil SR, Kimberling WJ, et al. 1977. Racial differences in the frequency оf Q- and С-chromosomal heteromorphism. Nature. 268: 631-632.
View at Publisher | View at Google Scholar - Yamada К. and Hasegawa Т. 1978. Types and frequencies of Q-variant chromosomes in а Japanese population. Нum. Genet., 44: 89-98.
View at Publisher | View at Google Scholar - Al-Nassar КЕ, Palmer СG, Connealy PМ, Рао-Lo Yu. 1981. The genetic structure of the Kuwaiti population. II. The distribution of Q-band chromosomal heteromorphisms. Нum Genet. 57: 423-427.
View at Publisher | View at Google Scholar - Ibraimov АI. 1993. The origin of modern humans: а cytogenetic model. Нum Evol. 8(2): 81-91.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ. 1982a. Human chromosomal polymorphism. II. Chromosomal С-polymorphism in Mongoloid populations of Central Asia. Hum Genet. 60: 8-9.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ. 1982b. Human chromosomal polymorphism. III. Chromosomal Q-polymorphism in Mongoloids of Northern Asia. Hum Genet. 62: 252-257.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ. 1982c. Human chromosomal polymorphism. IV. Q-polymorphism in Russians living in Kirghizia. Hum Genet. 62: 258-260.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ. 1982d. Human chromosomal polymorphism. V. Chromosomal Q-polymorphism in African populations. Hum Genet. 62: 261-265.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov М М. 1985. Q-band polymorphism in the autosomes and the Y chromosome in human populations. In: Progress and Topics in Cytogenetics. The Y chromosome. Part А. Basic characteristics of Y chromosome. А. А. Sandberg (Ed). Alan R. Liss, Inc., New York. USA, pp. 213-287.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ, Axenrod ЕI, Kurmanova G.U. 1986. Human chromosomal polymorphism. IX. Further data on the possible selective value of chromosomal Q-heterochromatin material. Hum Genet. 73: 151-156.
View at Publisher | View at Google Scholar - Ibraimov АI, Kurmanova GU, Ginsburg ЕК, Aksenovich TI, Axenrod ЕI. 1990. Chromosomal Q-heterochromatin regions in native highlanders of Pamir and Tien-Shan and in newcomers. Cytobios. 63: 71-82.
View at Publisher | View at Google Scholar - Ibraimov АI, Axenrod ЕI, Kurmanova GU, Turapov ОА. 1991. Chromosomal Q-heterochromatin regions in the indigenous population of the Northern part of West Siberia and in new migrants. Cytobios. 67: 95-100.
View at Publisher | View at Google Scholar - Ibraimov AI, Karagulova GO, Kim EY. 1997. Chromosomal Q-heterochromatin regions in indigenous populations of the Northern India. Ind. J. Hum. Genet. 3: 77-81.
View at Publisher | View at Google Scholar - Ibraimov AI, Akanov AA, Meymanaliev TS, Karakushukova AS, Kudrina NO, Sharipov KO, Smailova RD. 2013. Chromosomal Q-heterochromatin polymorphisms in 3 ethnic groups (Kazakhs, Russians and Uygurs) of Kazakhstan. Int J Genet. 5(1): 121-124.
View at Publisher | View at Google Scholar - Kalz L, Kalz-Fuller B, Hedge S, Schwanitz G. 2005. Polymorphism of Q-band heterochromatin; qualitative and quantitative analyses of features in 3 ethnic groups (Europeans, Indians, and Turks). Int J Hum Genet. 5(2): 153-163.
View at Publisher | View at Google Scholar - Décsey K, Bellovits O, Bujdoso GM. 2006. Human chromosomal polymorphism in Hungarian sample. Int J Hum Genet. 6(3): 77-183.
View at Publisher | View at Google Scholar - Caspersson Т, Zech L, Johansson С. 1970. Differential binding of alkilating fluorochromes in human chromosomes. Ехр Cell Res. 60: 315-319.
View at Publisher | View at Google Scholar - Arrighi FЕ, Hsu ТС. 1971. Localization of heterochromatin in human chromosomes. Cytogenetics. 10: 81-86.
View at Publisher | View at Google Scholar - Paris Conference, 1971, and Supplement 1975. Standartization in human cytogenetics. Birth Defects: Original Article Series, XI, 1-84. The National Foundation, New York.
View at Publisher | View at Google Scholar - Pearson PL. 1973. The uniqueness of the human karyotype. In: Chromosome identification techniques and application in biology and medicine. Caspersson Т. and Zech L. (eds). New York, London. Academic Press, p. 145.
View at Publisher | View at Google Scholar - Pearson PL. 1977. Pattern of bands, polymorphism and evolution of primates. In Molecular structure of human chromosomes. Yunis J.J. (Ed). Acad. Press. p. 267.
View at Publisher | View at Google Scholar - Chiarelli B, Lin CC. 1972. Comparison of florescence patterns in human and chimpanzee chromosomes. Genet Phaenen 15: 103-106.
View at Publisher | View at Google Scholar - Ibraimov A.I. 2020b. Chromosomal Q-heterochromatin in the Human Genome. Cambridge Scholars Publishing.
View at Publisher | View at Google Scholar - Ibraimov AI. 2018a. Why do not all people Ill with High-Altitude Pulmonary Edema? J Cardiol Diagnost Res. 1(1): 13-18.
View at Publisher | View at Google Scholar - Erdtmann В. 1982. Aspects of evaluation, significance, and evolution of human С-band heteromorphism. Нum Genet. 61: 281-294.
View at Publisher | View at Google Scholar - Ibraimov AI. 2004. The origin of condensed chromatin, cell thermoregulation and multicellularity. Complexus. 2: 23-34.
View at Publisher | View at Google Scholar - Ibraimov AI, Tabaldiev SK. 2007. Condensed chromatin, cell thermoregulation and human body heat conductivity. J Hum Ecol. 21(1): 1-22.
View at Publisher | View at Google Scholar - Ibraimov AI. 2016a. Chromosomal Q-Heterochromatin Polymorphism in Patients with Alimentary Obesity. Biol. Med. (Aligarh). 8: 275. DOI: 10.4172/0974-8369.1000275.
View at Publisher | View at Google Scholar - Ibraimov AI. 2016b. Chromosomal Q-heterochromatin Regions in Alcoholics and Drug Addicts. Biol. Med. (Aligarh). 8: 346. DOI: 10.4172/0974-8369.1000346.
View at Publisher | View at Google Scholar - Ibraimov AI. 2017a. Cell Thermoregulation: Problems, Advances and Perspectives. J Mol Biol Res. 7(1): 58-79. doi:10.5539/jmbr.v7n1p58.
View at Publisher | View at Google Scholar - Ibraimov AI. 2017b. From 48 to 46 chromosomes: Origin of Man. J Mol Biol Res. 7(l): 80-87.
View at Publisher | View at Google Scholar - Ibraimov AI. 2018b. Human Body Heat Conductivity in norm and pathology: A review. Advance Research Journal of Multidisciplinary Discoveries. 32(3): 12-21.
View at Publisher | View at Google Scholar - Ibraimov AI. 2019a. Chromosomal Q-heterochromatin Polymorphism and Sport. Int J Biol Med. 1: 45-52.
View at Publisher | View at Google Scholar - Ibraimov AI. 2019b. Cell thermoregulation and origin of homeothermic animals, Current Research in Biochemistry and Molecular Biology, 1(1): 10-13. http://dx.doi.org/10.33702/crbmb.2019.1.1.3.
View at Publisher | View at Google Scholar - Ibraimov AI. 2020a. The origin of the human karyotype: its uniqueness, causes and effects, Current Research in Biochemistry and Molecular Biology, 1, 1: 9-20.
View at Publisher | View at Google Scholar - Ibraimov AI, Akanov AA, Meimanaliev TS, Sharipov KO, Smailova RD, Dosymbekova R. 2014b. Human Chromosomal Q-heterochromatin Polymorphism and Its Relation to Body Heat Conductivity. Int J Genet. 6(1): 142-148.
View at Publisher | View at Google Scholar - Ibraimov AI. 2018. Chromocenters and Cell Thermoregulation. J Biol Med Res. Vol.2 No.3:19.
View at Publisher | View at Google Scholar - Ibraimov AI. 2020c. Chromosome Bands and Cell Thermoregulation. Current Research in Biochemistry and Molecular Biology. 2(1), http://dx.doi.org/10.33702.
View at Publisher | View at Google Scholar - Albrecht-Buehler G. 1990. In defense of “nonmolecular” cell biology. Inter Rev Cytol, 120: 191-241.
View at Publisher | View at Google Scholar - Schmid М, Vogel W, Krone W. 1975. Attraction between centric heterochromatin of human chromosomes. Cytogenet. Cell Genet. 15: 66-80.
View at Publisher | View at Google Scholar - Schöfer C, Weipoltshammer K. 2018. Nucleolus and chromatin. Histochem Cell Biol, 150(3) : 209-225. https://doi.org/10.1007/s00418-018-1696-3.
View at Publisher | View at Google Scholar - Thermoregulation. 1997. С.М. Blatties. (Ed). Ann. New York Acad. Sci., vol. 813.
View at Publisher | View at Google Scholar - Romanovsky AA. 2007. Thermoregulation: some concepts have changed. Functional architecture of the thermoregulatory system. Amer. J. Physiol.-Regul., Integr. Comp Physiol., 292, R37-R46.
View at Publisher | View at Google Scholar - Ibraimov АI, Mirrakhimov ММ, Nazarenko SА, Axenrod ЕI, Akbanova GА. 1982. Нuman chromosomal polymorphism. I. Chromosomal Q-polymorphism in Mongoloid populations of Central Asia. Hum Genet. 60: 1-7.
View at Publisher | View at Google Scholar - Ibraimov AI, Akanov AA, Meymanaliev TS, Smailova RD, Baygazieva GD. 2014a. Chromosomal Q-heterochromatin and age in human population. J Mol Biol Res. 4(1): 1-9.
View at Publisher | View at Google Scholar - Ibraimov AI, Karagulova GO. 2006. Chromosomal Q-heterochromatin regions in individuals of various age groups. Int J Hum Genet. 6(3): 219-228.
View at Publisher | View at Google Scholar
