

Research article | DOI: https://doi.org/10.31579/2835-7949/014
Islet Function and Insulin Secretion
- Rehan Haider 1*
- Asghar Mehdi
- Anjum Zehra
- Geetha Kumari Das
- Zameer Ahmed
1Riggs Pharmaceuticals Department of Pharmacy. UniversityofKarachi -Pakistan.
2Head Department of Pharmacology Fazaia Ruth Pfau Medical College Air University Karachi Pakistan.
3Assistant Professor Department of Microbiology University of Karachi Pakistan.
4GD Pharmaceutical Inc OPJS University Rajasthan India.
5Assistant Professor Dow University of Health Sciences Karachi Pakistan.
*Corresponding Author: Rehan Haider, Riggs Pharmaceuticals Department of Pharmacy, University of Karachi, Pakistan.
Citation: Rehan Haider, Asghar Mehdi, Geetha Kumari Das, Zameer Ahmed., Anjum Zehra., (2024), Islet Function and Insulin Secretion, Journal of Clinical Chemistry., 3(2); DOI: 10.31579/2835-7949/014
Copyright: © 2024, Rehan Haider. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 06 March 2024 | Accepted: 15 March 2024 | Published: 28 March 2024
Keywords: Islet function; insulin secretion; glucose homeostasis; beta cells; hormones; glucose metabolism; metabolic disorders
Abstract
The German anatomy student Paul Langerhans first described in 1869 the " islands of clear cells " distributed throughout the pancreas [1], but he did not realize the physiological significance of these cell clusters, which are today known as islets of Langerhans. We now know that Islets are the endocrine compartment of the pancreas, comprising approximately 2 – 3% of the total pancreatic volume. Islets are approximately spherical, with an average diameter of 100 – 200 μ m, and a healthy human pancreas may contain up to a million individual islets, each having its complex anatomy, blood supply, and innervation Langerhans islets are clusters of specialized endocrine cells located in the pancreas. These islets play crucial roles in maintaining glucose homeostasis by regulating insulin secretion. Insulin, a hormone produced by beta cells within islets, is essential for the uptake and utilization of glucose by various tissues in the body. The function of islets and precise regulation of insulin secretion involves a complex interplay of multiple factors. Glucose is the primary regulator of insulin release. Elevated blood glucose levels stimulate beta cells to release insulin, which promotes glucose uptake by cells, particularly in the liver, muscle, and adipose tissues. Insulin secretion is tightly regulated through a feedback mechanism, which ensures that insulin release matches the body's metabolic demands.
Introduction
Islet s structure and function Islet anatomy. A typical mammalian islet comprises several thousand endocrine cells, including insulin-expressing β- cells ( ∼ 60% of adult human islet cells), glucagon-expressing α- cells (20 – 30%), somatostatin-expressing δ-cells ( ∼ 10%), pancreatic polypeptide-expressing cells ( < 5>
Intra - Iset interactions
The anatomic organization of the islet has a profound influence on the ability of β- cells to recognize and respond to physiological signals [7 – 9]. There are several mechanisms through which islet cells can communicate, although the relative importance of these different mechanisms remains uncertain [10]. Islet cells are functionally coupled through a network of gap junctions, and gene deletion studies in mice have highlighted the importance of gap junctional coupling via connexin 36 in the regulation of insulin secretory responses [11,12]. Cell-to-cell contact through cell Surface adhesion molecules offer an alternative communication mechanism, and interactions mediated by E-cadherin [13,14] or ephrins [15] have been implicated in the regulation of β-cell function. A further level of control can be exerted via intra-islet paracrine and autocrine effects, in which a biologically active substance released by one islet cell can influence the functional status of a neighboring cell (paracrine) or itself (autocrine). Figure 6.2 shows some of the molecules that have been implicated in this type of intra-islet cell-to-cell communication. Thus, islet cells can interact with each other via the classic islet hormones – insulin, glucagon, and somatostatin [16 – 19]; via other products secreted by the endocrine cells, such as neurotransmitters or adenine nucleotides and divalent cations that are co-released with insulin [20 – 23]; and via other less well-known mechanisms, including the generation of gaseous signals such as nitric oxide and carbon monoxide [24 – 27]. The wide range of intra-islet interactions presumably reflects the requirement for fine-tuning and coordinating secretory responses of many individual islet cells to generate the rate and pattern of hormone secretion appropriate to the prevailing physiological conditions.
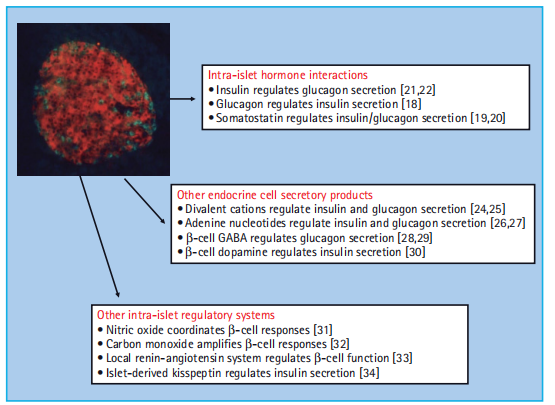
Figure 6:2 Intra - islet autocrine – paracrine interactions. The heterogeneous nature and complex anatomy of the islet enables numerous interactions between islet cells that are mediated by the release of biologically active molecules.
Insulin biosynthesis and storage
The ability to release insulin rapidly in response to metabolic demand, coupled with the relatively slow process of producing polypeptide hormones means that β - cells are highly specialized for the production and storage of insulin, to the extent that
Insulin comprises approximately 10% ( -10 pg/cell) of the total β-cell protein.
Biosynthesis of insulin In humans, the gene encoding preproinsulin, the precursor of insulin, is located on the short arm of chromosome 11 [28]. It is 1355 base pairs in length, and its coding region consists of three exons: the first encodes the signal peptide at the N-terminus of preproinsulin, the second the B chain and part of the C (connecting) peptide, and the third the rest of the C peptide and the A chain (Figure 6.3 ). Transcription and splicing to remove the sequences encoded by the introns yield a messenger RNA of 600 nucleotides, the translation of which gives rise to preproinsulin, an 11.5 - kDa polypeptide. The cellular processes and approximate timescales involved in insulin biosynthesis, processing, and storage are summarized in Figure 6.4.
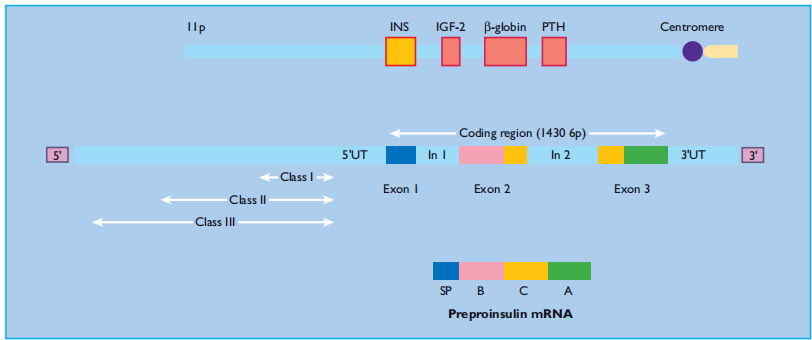
Figure 6:3 Structure of the human insulin gene. The coding region of the human insulin (INS) gene comprises three exons, which encode the signal peptide (SP), the B chain, C peptide and A chain. The exons are separated by two introns (In1 and In2). Beyond the 5 ′ untranslated region (5 ′ UT), upstream of the coding sequence, lies a hyper variable region in which three alleles (classes I, II and III) can be distinguished by their size.
Preproinsulin is rapidly ( < 1>
chains of insulin (21 and 30 amino acid residues, respectively) joined by the C-peptide (30 – 35 amino acids). The structural conformations of proinsulin and insulin are very similar, and a major function of the C peptide is to align the disulfide bridges that link the A and B chains such that the molecule is correctly folded for cleavage (Figure 6.5 ). Proinsulin is transported in micro vesicles to the Golgi apparatus, where it is packaged into membrane-bound vesicles, known as secretory granules. The conversion of proinsulin to insulin is initiated in the Golgi complex and continues.
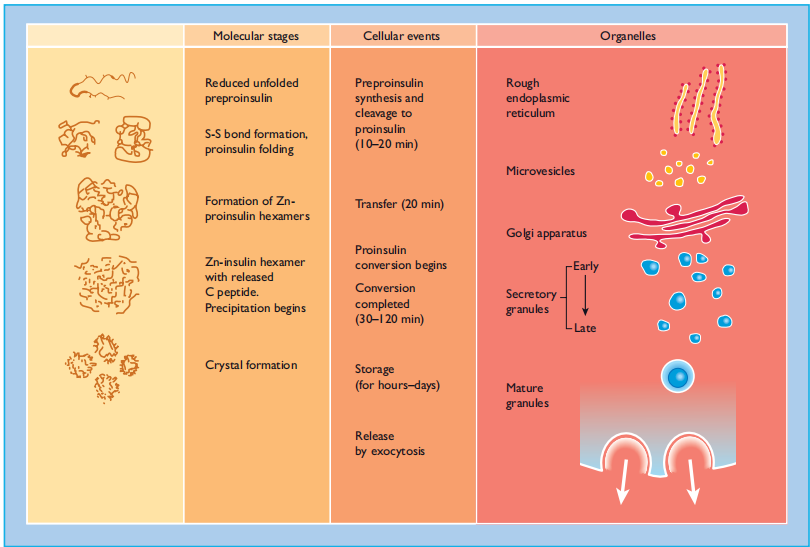
Figure 6:4 The intracellular pathways of (pro)insulin biosynthesis, processing and storage. The molecular folding of the proinsulin molecule, its conversion to insulin and the subsequent arrangement of the insulin hexamers into a regular pattern are shown at the left. The time course of the various processes, and the organelles involved are also shown.
within the maturing secretory granule through the sequential action of two endopeptidases (prohormone convertases 2 and 3) and carboxypeptidase H [29], which remove the C peptide chain, liberating two cleavage dipeptides and finally yielding insulin (Figure 6.5 ). Insulin and C peptides are stored together in secretory granules and are ultimately released in equimolar amounts by a process of regulated exocytosis. Under normal conditions, > 95% of the secreted product is insulin (and C-peptide), and < 5>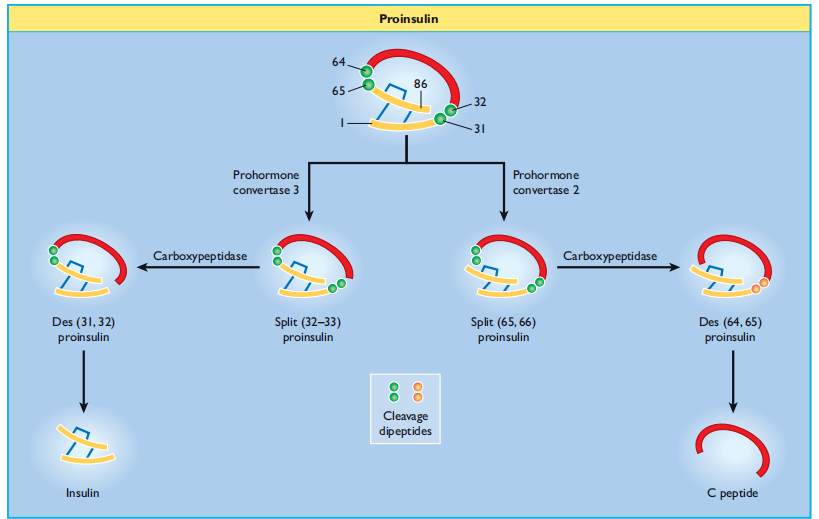
Figure 6:5 Insulin biosynthesis and processing. Proinsulin is cleaved on the C - terminal side of two dipeptides, namely Arg 31 – Arg 32 (by prohormone convertase 3) and Lys 64 – Arg 65 (prohormone convertase 2). The cleavage dipeptides are liberated, so yielding the “ split ” proinsulin products and ultimately insulin and C peptide.
β- cells respond to increases in the circulating concentrations of nutrients by increasing insulin production in addition to increasing insulin secretion, thus maintaining insulin stores [30]. Acute ( < 2>
Regulation of insulin s secretion
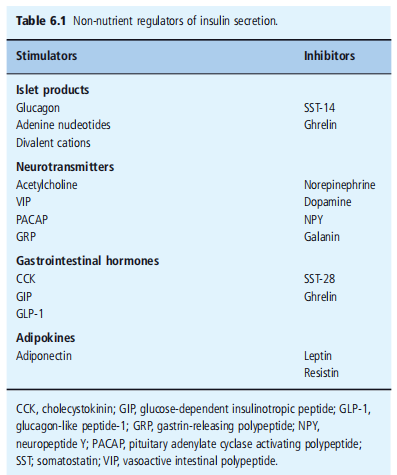
To ensure that circulating levels of insulin are appropriate for the prevailing metabolic status, β- cells are equipped with mechanisms to detect changes in circulating nutrients, hormone levels, and autonomic nervous system activity. Moreover, β - cells have fail-safe mechanisms for coordinating this afferent information and responding to the appropriate secretion of insulin. The major physiological determinant of insulin secretion in humans is the circulating concentration of glucose and other nutrients, including amino acids and fatty acids. These nutrients possess the ability to initiate an insulin secretory response, so When nutrients are absorbed from the gastrointestinal system, the β-cell detects changes in circulating nutrients and releases insulin to enable the uptake and metabolism or storage of nutrients by the target tissues. The consequent decrease in circulating nutrients is detected by the β - cells, which switch off Insulin secretion to prevent hypoglycemia. The responses of β- cells to nutrient initiators of insulin secretion can be modified by a variety of hormones and neurotransmitters that amplify or occasionally inhibit nutrient-induced responses (Table 6.1 ). Under normoglycemic conditions, these agents have little or there is no effect on insulin secretion, a mechanism that prevents inappropriate secretion of insulin, which would result in potentially harmful hypoglycemia. These agents are often referred to as potentiators of insulin secretion to distinguish them from the nutrients that initiate the secretory response. The overall insulin output depends on the relative input from initiators and potentiators at the level of individual β cells, synchronization of secretory activity between β - cells in individual islets, and coordination of secretion between hundreds of thousands of islets in the human.
pancreas. This section considers the mechanisms employed by β- cells to recognize and respond to nutrient initiators and non-nutrient potentiators of insulin secretion. Nutrient-induced insulin secretion Nutrient metabolism
Pancreatic β - cells respond to small changes in extracellular glucose concentrations within a narrow physiologic range and the mechanisms through which β - cells couple changes in nutrient metabolism to regulated exocytosis of insulin is becoming increasingly well-understood. Glucose is transported into β- cells via high-capacity glucose transporters ( GLUT2 in rodents and GLUT1, 2, and 3 in humans [47,48] ), enabling rapid equilibration of extracellular and intracellular glucose concentrations. Once inside the β-cell, glucose is phosphorylated by glucokinase, which acts as a " glucose sensor, " coupling insulin secretion to the prevailing glucose level [49]. The dose-response curve of glucose-induced insulin secretion from isolated islets is sigmoidal (Figure 6.6 ) and is primarily determined by glucokinase activity. Glucose concentrations below 5 mmol/L do not affect the rates of insulin release, and the rate of secretion increases progressively at extracellular glucose levels between 5 and ∼ 15 mmol/L, with half-maximal stimulation at ∼ 8 mmol/L. The time course of the insulin secretory response to elevated glucose levels is characterized by a rapidly rising but transient first phase, followed by a maintained and prolonged second phase, as shown in Figure 6.7. This profile of insulin secretion is obtained by measuring insulin levels following a glucose load in vivo, or by assessing the secretory output from the perfused pancreas or isolated islets, suggesting that the characteristic biphasic secretion pattern is an intrinsic property of the islets. ATP - sensitive potassium channels and membrane depolarization In the absence of extracellular glucose, the β-cell membrane potential is maintained close to the potassium equilibrium potential by the efflux of potassium ions through the inwardly rectifying potassium channels. These channels were called ATP - sensitive potassium (K ATP ) channels because the application of ATP to the The cytosolic surface of β-cell membrane patches results in rapid, reversible inhibition of resting membrane permeability to potassium ions [50]. This property of the K ATP channel is pivotal for linking glucose metabolism to insulin secretion. Thus, it is now established that ATP generation following glucose metabolism, in conjunction with the concomitant lowering of ADP levels, leads to the closure of β-cell K ATP channels. Channel closure and subsequent reduction in potassium efflux promote depolarization of the β - cell membrane and the influx of calcium ions through Voltage-dependent l-type calcium channels. The resultant increase in cytosolic Ca 2+ triggers the exocytosis of insulin secretory granules, thus initiating the insulin secretory response (Figure 6.8 ).
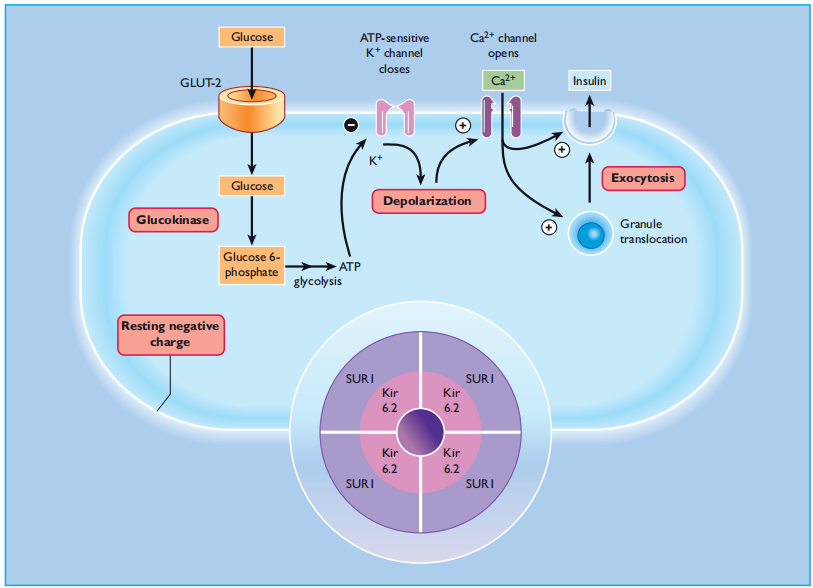
Figure 6:8 Intracellular mechanisms through which glucose stimulates insulin secretion. Glucose is metabolized within the β - cell to generate ATP, which closes ATP - sensitive potassium channels in the cell membrane. This prevents potassium ions from leaving the cell, causing membrane depolarization, which in turn opens voltage - gated calcium channels in the membrane and allows calcium ions to enter the cell. The increase in cytosolic calcium initiates granule exocytosis. Sulfonylureas act downstream of glucose metabolism, by binding to the SUR1 component of the K ATP channel (inset). GLUT, glucose transporter.
Around the time that the K ATP channels were established as the link between the metabolic and electrophysiological effects of glucose, they were also identified as the cellular target for sulfonylureas. The capacity of sulfonylureas to close K ATP channels explains their effectiveness in type 2 diabetes where the β - cells no longer respond adequately to glucose, as the usual pathway for coupling glucose metabolism with insulin secretion is bypassed. The β- cellular okay ATP channel is a hetero-octamer shaped from four potassium channel subunits ( Kir6.2) and four sulfonylurea receptor subunits (SUR1) [51]. The Kir6.2 subunits form the pore thru, where potassium ions flow and are surrounded by SUR1 subunits, which have a regulatory role (determine 6.8, inset). ATP and sulfonylureas result in channel closure by binding to the Kir6.2 and SUR1 subunits, respectively, at the same time ADP turns on the channels by binding to a nucleotide-binding area on the SUR1 subunit. Diazoxide, an inhibitor of insulin secretion, also binds to the SUR1 subunit to open channels. The imperative function of ok ATP channels in β-mobile glucose makes them obvious candidates for β-mobile dysfunction in type 2 diabetes. Earlier studies in patients with type 2 diabetes, adulthood-onset diabetes of the younger (MODY), or gestational diabetes, failed to hit upon any Kir6.2 gene mutations that compromised channel features [52,53]. Since then, larger-scale studies of variations in genes encoding Kir6.2 and SUR1 have demonstrated polymorphisms associated with an elevated risk of type 2 diabetes [54]. In addition, activating mutations within the Kir6.2 gene are causal for cases of everlasting neonatal diabetes (PNDM) [55], which has enabled insulin-structured PNDM sufferers to acquire glycemic control with sulfonylurea treatment on their own. In evaluation, lack of β - cell practical ok ATP channel pastime has been implicated in the pathogenesis of persistent Hyperinsulinemic hypoglycemia of infancy (PHHI) [56], a situation characterized by hypersecretion of insulin despite profound hypoglycemia. several mutations in both the Kir6.2 and SUR1 subunits had been identified in sufferers with PHHI and those are thought to be responsible for the excessive impairment of glucose homeostasis in these people [57].
Calcium and other intracellular effectors
Intracellular calcium is the most important effector of the nutrient-mediated insulin secretory reaction, linking depolarization with exocytosis of insulin secretory granules (figure 6.8 ). A massive electrochemical concentration gradient ( ∼10 000 - fold) of calcium is maintained across the β - cell plasma membrane via an aggregate of membrane-associated calcium extrusion systems and lively calcium sequestration within intracellular organelles. The major direction through which calcium is extended in β - cells are via an influx of extracellular calcium through the voltage-based calcium channels that open in reaction to β - cellular depolarization, and it has been expected that every β - mobile incorporates about 500 l - type channels [58]. Studies with permeabilized β- cells have confirmed that elevations in intracellular calcium alone are sufficient to provoke insulin secretion [59], and situations that elevate intracellular calcium normally stimulate insulin release. Growth in cytosolic calcium is vital for the initiation of insulin secretion through glucose and other nutrients: preventing calcium influx by removing extracellular calcium or by pharmacologic blockade of voltage-established calcium channels abolishes nutrient-induced insulin secretion. Glucose and different vitamins also set off calcium-based activation of β - cellular phospholipase C (%) [60] The generation of inositol 1,4,5 - trisphosphate (IP 3 ) and diacylglycerol (DAG), both of which serve second-messenger functions in β- cells [61]. The generation of IP 3 leads to the rapid mobilization of intracellular calcium, but the significance of this in secretory responses to nutrients is uncertain, and it is likely to
have little more than a modulatory role, amplifying the elevation in cytosolic calcium concentration induced by the influx of extracellular calcium. The elevations in intracellular calcium are transduced into the regulated secretion of insulin by intracellular calcium sensing systems within β- cells. Important among these are the calcium-dependent protein kinases, which include myosin light chain kinases, calcium/phospholipid-dependent kinases, and calcium/calmodulin-dependent kinases (CaMKs). CaMKs are protein kinases that are activated in the presence of calcium and the calcium-binding protein, calmodulin, and several Studies have implicated CaMKII in insulin secretory responses [61]. It has been proposed that CaMK II activation is responsible for the initiation of insulin secretion in response to glucose and other nutrients, and for enhancing nutrient-induced secretion in response to receptor agonists that elevate intracellular calcium [61] . Cytosolic PLA 2 (cPLA 2 ) is another β-cell calcium-sensitive enzyme. It is activated by the concentration of calcium that is achieved in stimulated β- cells and generates arachidonic acid (AA) by the hydrolysis of membrane phosphatidylcholine. AA is capable of stimulating insulin secretion in glucose- and calcium-independent manner, and it is further metabolized in islets by the cyclooxygenase (COX) pathways to produce prostaglandins and thromboxanes, and by the lipoxygenase (LOX) pathways to generate hydroperoxy eicosatetraenoic acids (HPETES), hydroxy eicosatetraenoic acids (HETES), and leukotrienes. The precise roles(s) of AA derivatives in the islet function remain uncertain because experimental investigations have relied on COX and LOX inhibitors of poor specificity, but earlier reports that prostaglandins are largely inhibitory in rodent islets [62] have not been supported by recent studies on human islets [63]. Calcium sensors are also important at the later stages of the secretory pathway, where the calcium-sensitive synaptotagmin proteins are involved in the formation of the exocytotic SNARE complex, as described above, confers calcium sensitivity on the initiation and rate of exocytotic release of insulin secretory granules. [44]. The elevation in intracellular calcium induced by nutrients activates other effector systems in β- cells, including PLC and PLA 2, as discussed above, and calcium-sensitive adenylate cyclase isoforms, which generate cyclic AMP from ATP. Although These signaling systems are undoubtedly important in the regulation of β - cells by non-nutrients, and their role in nutrient-induced insulin secretion is still uncertain. Thus, DAG generated by glucose-induced PLC activation has the potential to activate some Protein Kinase C (PKC) Isoforms. PKC was first identified as a calcium- and phospholipid-sensitive DAG-activated protein kinase, but it has become apparent that some isoforms of PKC require neither calcium nor DAG for activation. The isoforms are classified into three groups: calcium and DAG-sensitive (conventional), calcium-independent, DAG-sensitive (novel), calcium and DAG-independent (atypical), and β- cells contain conventional, atypical, and novel PKC isoforms [64]. The early literature on the role of PKC in nutrient-induced insulin secretion is confusing but several studies have shown that glucose-induced insulin secretion is maintained under conditions where DAG-sensitive PKC isoforms are depleted, suggesting that conventional and novel PKC isoforms are not required for insulin secretion in response to glucose [61,65]. The role of cAMP in the insulin secretory response to nutrients remains unclear. Cyclic AMP has the potential to influence insulin secretion by activating cyclic AMP-dependent protein kinase A (PKA) or via cyclic AMP-regulated guanine nucleotide exchange factors known as exchange proteins activated by cyclic AMP (EPACs) [66]. However, elevations in β - cell cyclic AMP do not stimulate insulin secretion at sub-stimulatory glucose concentrations, and the secretagogue The effects of glucose can be maintained in the presence of competitive antagonists of cAMP binding to PKA or EPACs [67]. These observations suggest that cyclic AMP does not act as the primary trigger of nutrient - stimulated β - cell secretory function, but more recent observations linking glucose-induced oscillations in β - cell cyclic AMP to oscillations in insulin secretion [68] suggest that a role for this messenger system in nutrient-induced Insulin secretion cannot be ruled out.
KATP channel - I independent pathways
Since the early reports linking K ATP channel closure to the exocytotic release of insulin, it has become apparent that β - cells also possess a K ATP channel-independent stimulus-secretion coupling pathway, which is termed the amplifying pathway to distinguish it from the triggering pathway that is activated by K ATP channel closure [69]. Studies in which β-cell calcium is elevated by depolarization and other ATP channels maintained within the open kingdom with the aid of diazoxide have indicated that glucose, at concentrations as low as 1 – 6 mmol/L, is still capable of stimulating insulin secretion [70]. The mechanisms by which glucose stimulates insulin secretion in an ATP-channel-independent manner have not yet been established [71]. However, glucose must be metabolized, and there may be convincing evidence that adjustments in adenine nucleotides are worrying [72], although it has been established that activation of PKA and % is not required. It has been suggested that okay ATP, the unbiased amplifying pathway, is impaired in type 2 diabetes and the identification of novel healing strategies targeted at this pathway may be beneficial in restoring β- cellular function in patients with type 2 diabetes [69].
Amino acids
Numerous amino acids stimulate insulin secretion, both in vivo and in vitro. Most require glucose, but some, including leucine, lysine, and arginine, can stimulate insulin secretion in the absence of glucose, and therefore qualify as initiators of secretion. Leucine enters islets by using a sodium-independent delivery system and stimulates a biphasic boom in insulin launches. The effects of leucine on β- cellular membrane potential, ion flux, and insulin secretion are much like but smaller than those of glucose [73]. For this reason, the metabolism of leucine inside β - cells decreases the potassium permeability town, inflicting depolarization and activation of L-type calcium channels through which calcium enters β - cells and initiates insulin secretion. Likewise, leucine is capable of setting off the amplifying pathway of insulin secretion in a k ATP channel-unbiased way, as defined above for glucose. The charged amino acids, lysine, and arginine, pass through the β-mobile plasma membrane via a transport device particular for cationic amino acids. It is generally believed that the accumulation of these positively charged molecules depolarizes the β- cellular membrane without delay, leading to calcium influx. Regulation of insulin secretion by non-nutrients The complex mechanisms that have developed to permit modifications in extracellular vitamins to provoke an exocytotic secretory reaction are limited to pancreatic β - cells, and possibly to a subset of hypothalamic neurons [74]. however, the mechanisms that β - cells use to understand and respond to non-nutrient potentiators of secretion are ubiquitous in mammalian cells, and so are included handiest in brief on this phase, followed via an evaluation of the physiologically relevant non-nutrient regulators of β - cellular features. maximum, if no longer all, non-nutrient modulators of insulin secretion impact the β - mobile via binding to and activating specific receptors on the extracellular surface. because of its crucial function in coordinating entire-frame fuel homeostasis the β - mobile expresses receptors for an extensive range of biologically active peptides, glycoproteins, and
neurotransmitters (Desk 6.1 ). However, receptor occupancy typically occurs within the activation of a confined quantity of in-travel mobile effector structures, which have been added inside the segment during the nutrient-induced insulin phase (Figures 6.9 and 6.10 ).
Islet hormones
There is convincing evidence of complex intra-islet interactions via molecules released from islet endocrine cells (Figure 6.2 ). The physiological relevance of some of these interactions is still uncertain, but some of the intra-islet factors that are thought to influence insulin secretion are discussed briefly in this section. It is now clear that β- cells express insulin receptors and the associated intracellular signaling elements, suggesting the existence of autocrine and/or paracrine feedback regulation of β-cell function [19,75]. Earlier suggestions that secreted insulin regulates insulin secretion [76] have not been confirmed [19,77], and the main feedback function of insulin on β - cells is to regulate β - cell gene expression [75,78] and β - cell mass through effects on proliferation and apoptosis [19,79] Glucagon is a 29 amino acid peptide secreted by the pancreatic islets. - cells. The precursor pro-glucagon undergoes differential translational processing in the gut to produce highly specific peptides with superior receptors and biological activity. These include glucagon-like peptide 1 (GLP-1) (7-36) amide, the 'incretin' hormone described below, and GLP-2, which promotes intestinal lining development. Glucagon secretion is regulated by nutrients, islets, gastrointestinal hormones, and the autonomic nervous system, with hypoglycemia and sympathetic Nervous input is an important stimulator of glucagon secretion [80]. Glucagon enhances insulin secretion through the stimulatory G-protein (G s )-coupled activation of adenylate cyclase and the consequent increase in intracellular cyclic AMP . Somatostatin (SST) is expressed by islet δ - cells and in numerous other sites, including the central nervous system and D cells of the gastrointestinal tract, where It acts predominantly as an inhibitor of endocrine and exocrine secretion [81]. Theprecursor, pro somatostatin, is processed by alternative pathways: in islets and the CNS SST - 14 is generated, and SST - 28, the major circulating form of SST in humans is produced in the gastrointestinal tract [82]. SST secretion is regulated by a variety of nutrient, endocrine, and neural factors [17,81,83]. Islets express five different somatostatin receptor (SSTR) subtypes, and SST-14 released from islet δ - cells havea tonic inhibitory effect on insulin and glucagon secretion [17], via activation of SSTR5, and SSTR2, respectively [84,85]. SST receptors are coupled via an inhibitory G - protein(s) inhibit adenylate cyclase and decrease the formation of cyclic AMP [86] and ion channels that cause hyper polarization of the β-cell membrane and reductions in intracellular calcium [87]. Pancreatic polypeptide (PP) is a 36 amino acid peptide produced by PP cells found in the mantle of islets, predominantly those located in the head of the pancreas. PP secretion is mainly regulated via cholinergic parasympathetic stimulation [81], but the physiological function of PP as a circulating hormone or as an intra-islet signal remains uncertain [88]. Ghrelin is a 23 amino acid peptide first identify ed in the gastrointestinal system but is now known to also be expressed in islets ε - cells, which are localized to the islet mantle in rodents, and which appear to be developmentally distinct from the classic islet endocrine cells [89,90]. The physiological function of ε-cell-derived ghrelin has not yet been established, but most experimental evidence suggests an inhibitory role in the regulation of insulin secretion [91], analogous to that of δ-cell SST [17].
Neural control of insulin s secretion
The association of nerve fibers with islets was shown over 100 years ago by silver staining techniques [92]; since then, it has become well-established that islets are innervated by cholinergic, adrenergic, and peptidergic autonomic nerves.
Parasympathetic (cholinergic) fibers originate in the dorsal motor nucleus of the vagus, and sympathetic (adrenergic) fibers from the greater and middle splanchnic nerves penetrate the pancreas and terminate close to islet cells. Autonomic innervation of islets is important in regulating insulin secretion, with enhanced insulin output following activation of parasympathetic nerves and decreased insulin secretion in response to increased sympathetic activity. The autonomic nervous regulation of islet hormone secretion is thought to be involved in the The cephalic phase of insulin secretion during feeding synchronizes islets to generate oscillations of hormone secretion and regulates islet secretory responses to metabolic stress [5].
Neurotransmitters: acetylcholine and norepinephrine
The numerous parasympathetic nerve fibers that innervate islets are post-ganglionic and originate from the intra-pancreatic ganglia, which are controlled by pre-ganglionic fibers originating from the dorsal vagal nucleus [5]. Acetylcholine is the major post ganglionic parasympathetic neurotransmitter, which stimulates the release of insulin and glucagon in a variety of mammalian species [5,93,94]. In β - cells, acetylcholine acts predominantly via M3 receptors [93,95] to activate PLC (Figure 6.10 ), generating IP 3 and DAG, which act to amplify the effects of glucose by elevating cytosolic calcium levels and activating PKC [61]. Activation of β - cell muscarinic receptors can also lead to the activation of PLA 2, with the subsequent generation of AA and lysophosphatidylcholine, which can further enhance nutrient-induced insulin secretion. Acetylcholine also depolarizes the plasma membrane by affecting Na + conductivity, and this additional depolarization induces sustained increases in cytosolic calcium [93].
Islets also receive extensive sympathetic innervation from post-ganglionic nerves, whose cell bodies are located in the celiac or paravertebral ganglia, while the pre-ganglionic nerves originate from the hypothalamus [5]. The major sympathetic neurotransmitter, norepinephrine (noradrenaline), can exert both positive and negative effects on hormone secretion. Thus, norepinephrine can exert direct stimulatory effects on the β - cell via β2 - adrenoreceptors [96], or inhibitory effects via α2 - adrenoreceptors [97], and the net effect of norepinephrine may depend on the Relative expression levels of these receptor subtypes. Differences in the expression levels of adrenoreceptor subtypes between species probably account for the differential effects of β- adrenergic agonists on human islets, where they are stimulatory, and rodent islets, where they are ineffective [98]. The stimulatory effects mediated by β2 - receptors occur by activation of adenylate cyclase and an increase in intracellular cyclic AMP (Figure 6.9 ) [99], while the inhibitory effect of α2 - receptor activation involves reductions in cyclic AMP and cytosolic calcium [86,100], and an unidentified inhibitory action at a more distal point in the Stimulus–secretion coupling mechanism [101]. In contrast, norepinephrine has direct stimulatory effects on glucagon secretion from α- cells mediated by both β2 and α2 receptor subtypes [5]. Circulating catecholamines secreted by the adrenal medulla (mainly epinephrine) also have the potential to influence islet hormone secretion through interactions with adrenoreceptors expressed on α and β cells.
Neuropeptides
Parasympathetic nerve fibers in islets contain several biologically active neuro peptides, including VIP, PACAP, and gastrin-releasing polypeptides (GRP), all of which are released by vagal activation and all of which stimulate the release of insulin and glucagon.VIP (28 amino acids) and PACAP (27 or 38 amino acids) are abundantly expressed neuro peptides that are broadly allotted in the parasympathetic nerves that supply the islets and gastrointestinal tract [6,102]. VIP and PACAP have comparable structures, and VIP1 and VIP2 receptors also have an affinity for PACAP. The Stimulatory outcomes of VIP and PACAP on insulin secretion in vitro and in vivo are through β- cellular VIP2 and p.c. receptors, respectively, and are thought to be involved in increases in intracellular cAMP (figure 6. nine ) and cytosolic calcium [6,103,104]. GRP is a 27 amino acid peptide that still stimulates the secretion of Insulin, glucagon, SST, and PP [5,94]. These outcomes of GRP are mediated through specific receptors and involve the activation of percent and the technology of IP three and DAG (determine 6.10 ) [102,105]. Sympathetic nerves comprise special Neuropeptides to the parasympathetic nerves, and these encompass NPY and galanin, both of which have inhibitory movements within the islets. NPY (36 amino acids) and galanin (29 amino acids) are expressed in fibers innervating each of the endocrine and exocrine pancreas [5,106]. each neuro-peptide inhibits basal and glucose-stimulated insulin secretion [86,106,107], even though variations among species have been suggested. each NPY and galanin act via specific c G i - coupled receptors to inhibit adenylate cyclase [108,109], and galanin may additionally have extra inhibitory consequences at an undefined ned overdue stage of exocytosis [86] Incretin and an adipokine regulation of I insulin s secretion It has been known for over 40 years that insulin secretion from islets is greater following oral rather than intravenous administration of glucose [110], and it is now known that this enhanced insulin secretory output is a consequence of the release of gastrointestinal-derived " incretin " hormones [111]. The main incretins implicated in elevated insulin response to absorbed nutrients after food intake are glucagon-like peptide 1 (GLP - 1), glucose-dependent insulinotropic peptide (GIP), and cholecystokinin (CCK), all of which are hormones secreted by specialized endocrine cells in the gastrointestinal tract in response to the absorption of nutrients [111,112]. These hormones are carried to islets in the blood and interact with specific receptors on the β-cell surface to stimulate insulin secretion.
Glucagon-like peptide 1
After food intake, intestinal L-cells secrete GLP - 1 in response to elevated levels of nutrients derived from carbohydrates, lipids, and proteins in the intestinal lumen [112,113]. GLP - 1 is generated by prohormone convertase 1 – 3 cleavage of proglucagon in the L - cells and is highly conserved in mammals, with identical amino acid sequences in humans and mice [82,113]. GLP - 1 is degraded by dipeptidyl protease 4 (DPP - 4), which cleaves two amino acids from the N-terminus. Full-length GLP - 1 (1 – 37) does not show biological activity, but the truncated peptides GLP - 1 (7 – 36) amide and GLP - 1 (7 – 37) are potent stimulators of insulin secretion, both in vitro and in vivo [114]. Observations that infusion of the peptide into individuals with type 2 diabetes before food intake improved insulin output and reduced the post-prandial increase in circulating glucose led to studies to determine whether GLP-1 and related peptides may be useful as therapies for type 2 diabetes. Reports of other beneficial effects of GLP-1, including its capacity to inhibit glucagon secretion, delay gastric emptying, and decrease food intake, have indicated its positive effects on normalizing postprandial glycemia, but its half-life of less than 2 min precludes its use as a diabetes therapy. Nonetheless, exenatide, a synthetic version of a GLP - 1 analog present in the saliva of the Gila monster lizard, has been developed for clinical use in type 2 diabetes [115]. It has approximately 50% amino acid homology with GLP - 1 thus allowing it to exert the same effects on islets as native GLP - 1, but its resistance to degradation by DPP-4 increases its half-life to approximately 2 h in vivo, which ensures effective regulation of blood glucose levels. Another GLP-1 analog, liraglutide, has a greatly extended half-life ( > 12
h) because of the incorporation of the fatty acid palmitate into the GLP - 1 sequence, allowing it to bind to plasma albumin and reduce its exposure to DPP - 4. Selective DPP-4 inhibitors such as sitagliptin are used clinically to normalize blood glucose levels in type 2 diabetes by extending the half-life of endogenous GLP - 1. GLP - 1, exenatide, and liraglutide act on specific G protein-coupled receptors [111] that are linked via G s to adenylate cyclase activation, which ultimately increases insulin secretion (Figure 6.9 ) [114]. Although cyclic AMP has been implicated In the majority of effects of GLP - 1 in islets, it may also close rat β-cell K ATP channels in a cyclic AMP-independent manner [116].
Glucose-dependent I insulinotropic p peptide
Glucose-dependent insulinotropic peptide (GIP), a 42 amino acid peptide, is released from K - cells in the duodenum and jejunum in response to the absorption of glucose, other actively transported sugars, amino acids, and long-chain fatty acids [112]. It was originally called " gastric inhibitory polypeptide " because of its inhibitory effects on acid secretion in the stomach, but its main physiological effects are now known to be the stimulation of insulin secretion in a glucose-dependent manner [102,111]. Like GLP - 1, GIP binds to G s - coupled receptors on the β - cell plasma membrane, with essentially the same downstream cascades leading to stimulation of insulin secretion (Figure 6.9 ) [102,111]. GIP is also reported to increase insulin release through the generation of AA via phospholipase A 2 activation [117] and closure of K ATP channels [111]. Although GLP-1 and GIP both enhance insulin output following their release in response to food intake, it seems unlikely that GIP-related peptides will be developed as therapies for type 2 diabetes because GIP stimulates glucagon secretion and inhibits GLP-1 release, and its infusion in individuals with type 2 diabetes is reported to worsen post-prandial hyperglycemia [118].
Cholecystokinin
CCK is another incretin hormone released from cells in the gastrointestinal tract in response to elevated fat and protein levels [112]. It was originally isolated f the porcine intestine as a 33 amino acid peptide, and the truncated CCK - 8 form stimulates insulin secretion in vitro and in vivo [119]. CCK - 8 acts at specific G q - coupled receptors on β - cells to activate PLC (Figure 6.10 )
, and potentiation of insulin secretion is completely dependent on PKC activation [120]. However, the physiological role of CCK as an incretin has not been established because high concentrations are required for its effects on insulin secretion, and its major function may be digestion in the duodenum.
Adipokines
obesity is a chance element for diabetes, and hormones (adipokines) launched from fats depots have been implicated in insulin resistance related to weight problems and type 2 diabetes [121]. a few adipokines inclusive of leptin, resistin, and adiponectin are also suggested to steer the islet feature. consequently, β - cells explicit Ob - Rb leptin receptors which, when activated through leptin, lead to the inhibition of insulin secretion [122], and precise deletion of β - mobile Ob - Rb receptors are associated with improved insulin secretion [123]. The inhibitory effects of leptin on glucose-stimulated insulin secretion have been attributed to the activation of β-cellular okay ATP channels [124] or c-Jun N-terminal kinases (JNKs) [125]. Leptin can also in addition impair β - mobile characteristics thru reductions in β - mobile mass [123,125]. Resistin, another adipocyte polypeptide, also inhibits glucose-stimulated insulin launch [126] and stimulates apoptosis of rat β- cells [127], suggesting that its features are comparable to those of leptin. However, resistin is not considered to be a real adipokine because, although it is secreted by excessive tiers from mouse adipocytes, it is not produced with the aid of human adipocytes, and excessive plasma resistin levels do not correlate with reduced insulin sensitivity [128]. However, resistin may have paracrine effects on β- mobile function in humans, as has been identified in human islets [129]. In contrast to leptin and resistin, adiponectin has defensive effects by improving insulin sensitivity, and reduced plasma adiponectin levels may also contribute to the development of type 2 diabetes [130]. The practical outcomes of adiponectin in β - cells have not been completely installed, however human and rat β- cells have explicit adiponectin receptors [131,132]. Adiponectin stimulates insulin secretion [131,132] and defends against β- mobile apoptosis [133]. The signaling cascades that couple adiponectin receptors to downstream effects in β cells are currently unknown.
Research Method
Diverse techniques can be employed to analyze islet function and insulin secretion. Some common techniques encompass in vitro experiments using removed islets or β- cells, in vivo studies using animal models, and clinical investigations involving human participants.
Result
Research on islet features and insulin secretion has provided insights into the mechanisms underlying glucose homeostasis and the pathogenesis of diabetes. Research has identified key elements related to insulin secretion, including ATP-touch potassium (KATP) channels, voltage-gated calcium channels, and various signaling pathways.
it's been determined that glucose plays a crucial role in regulating insulin secretion. When glucose tiers upward push, it enters β- cells via glucose transporters (GLUT2 in rodents, GLUT1 in humans), undergoes glycolysis, and results in extended ATP manufacturing. An elevated ATP/ADP ratio results in the closure of KATP channels, depolarization of the β-cellular membrane, and subsequent commencing of voltage-gated calcium channels. Calcium influx triggers insulin granule exocytosis and is released into the bloodstream. In addition to glucose, other factors can modulate islet characteristics and insulin secretion. These include incretin hormones (along with glucagon-like peptide-1 or GLP-1), neurotransmitters (e.g., acetylcholine), hormones (e.g., glucagon and somatostatin), and other metabolites. The interaction of these elements determines the overall reaction of pancreatic islets to adjustments in glucose titers. Various pathophysiological conditions can affect islet characteristics and insulin secretion. Type 1 diabetes is characterized by autoimmune destruction of β- cells, leading to an absolute deficiency of insulin. In type 2 diabetes, β- cellular dysfunction and insulin resistance contribute to impaired insulin secretion. Genetic elements, environmental elements (including food plan and lifestyle), and Epigenetic changes can affect islet characteristics and insulin secretions.
Discussion
This study's findings on islet characteristics and insulin secretion have extensive implications for our expertise in diabetes and the improvement of therapeutic strategies. By elucidating the molecular mechanisms involved in insulin secretion, researchers have recognized capability targets for drug interventions aimed at enhancing insulin launch or β-mobile survival.
moreover, the take a look at islet function has highlighted the complexity of the regulatory community governing insulin secretion. The interplay between glucose and various factors, including hormones and neurotransmitters, underscores the need for comprehensive expertise in these interactions to develop effective remedies for diabetes. Furthermore, research on islet function and insulin secretion has underscored the significance of individual variations in response to glucose and other modulating factors. Genetic and Epigenetic elements can contribute to inter-male or female variations in insulin secretion, which can also explain variations in the danger and progression of diabetes among people. Research on islet features and insulin secretion has deepened our understanding of the intricate mechanisms that underlie glucose homeostasis and diabetes. Research in this area holds promise for the development of novel healing methods and personalized interventions for diabetes control.
Conclusions
The pancreatic islets of Langerhans are complex endocrine organs containing several different types of endocrine cells, with extensive vasculature and autonomic nerve supply. Interactions between islet cells, the autonomic nervous system, and hormones secreted by the gastrointestinal system and adipose tissue enableth the appropriate release of islet hormones to regulate metabolic fuel usage and storage. The insulin-secreting β- cells within islets respond to changes in circulating nutrients by linking changes in nutrient metabolism to β-cell depolarization and calcium-dependent exocyst to release stored insulin. β - Cells can also respond to a wide range of hormones and neurotransmitters through conventional cell surface receptors that are linked to a variety of intracellular effectors systems regulating the release of insulin. The ability to detect nutrient, hormonal, and neural signals allows β- cells to integrate information about the prevailing metabolic status and to secrete insulin as required for glucose homeostasis. This detailed understanding of islet cell biology has led to the development of new treatments for type 2 diabetes, as evidenced by the recent introduction of GLP - 1agonistst and DPP - 4 inhibitors. Recent studies have suggested associations between type 2 diabetes and polymorphisms in genes associated with β-cell development or function; therefore, the current understanding of normal β-cell function may assist in identifying β-cell pathologies in type 2 diabetes.
Acknowledgment
The completion of this research project would not have been possible without the contributions and support of many individuals and organizations. We are deeply grateful to all those who played a role in the success of this project We also thank My Mentor [. Naweed Imam Syed Prof. Department of Cell Biology at the University of Calgary and Dr. Sadaf Ahmed Psychophysiology Lab University of Karachi for their invaluable input and support throughout this research. Their insights and expertise were instrumental in shaping the direction of this project.
Declaration of Interest
I at this moment declare that:
I have no pecuniary or other personal interest, direct or indirect, in any matter that raises or may raise a conflict with my duties as a manager of my office Management
Conflicts of Interest
The authors declare that they have no conflict of interest.
Financial support and sponsorship
No Funding
References
- Langerhans P . Beitrage zur mikroskopischen Anatomie der Bauchs - peicheldruse. Inaugural dissertation. Gustav Lange, ed. 1869 .
View at Publisher | View at Google Scholar - Cabrera O , Berman DM , Kenyon NS , Ricordi C , Berggren PO , Caicedo A . The unique cyto architecture of human pancreatic islets has implications for islet cell function. Proc Natl Acad Sci U S A 2006; 103: 2334 – 2339.
View at Publisher | View at Google Scholar - Bonner - Weir S . Anatomy of the islet of Langerhans . In: Samols E , ed. The Endocrine Pancreas . New York : Raven Press , 1991 ; 15 – 27 .
View at Publisher | View at Google Scholar - Nyman LR, Wells KS, Head WS, McCaughey M, Ford E, Brissova M, et al. Real-time, multidimensional in vivo imaging is used to investigate blood flow in mouse pancreatic islets. J Clin Invest 2008; 118: 3790 – 3797.
View at Publisher | View at Google Scholar - Ahren B . Autonomic regulation of islet hormone secretion: implications for health and disease. Diabetologia 2000 ; 43 : 393 – 410 .
View at Publisher | View at Google Scholar - Winzell MS , Ahren B . Role of VIP and PACAP in islet function.Peptides 2007; 28: 1805 – 1813.
View at Publisher | View at Google Scholar - Halban PA , Wollheim CB , Blondel B , Meda P , Niesor EN , Mintz DH. The possible importance of contact between pancreatic islet cells for the control of insulin release. Endocrinology 1982; 111: 86 – 94.
View at Publisher | View at Google Scholar - Halban PA, Powers SL, George KL, Bonner - Weir S . Spontaneous re-association of dispersed adult rat pancreatic islet cells into aggregates with three-dimensional architecture typical of native islets. Diabetes 1987; 36: 783 – 790.
View at Publisher | View at Google Scholar - Hop Croft DW, Mason DR, Scott RS . Insulin secretion from perifused rat pancreatic pseudo islets. In Vitro Cell Dev Biol 1985; 21: 421 – 427.
View at Publisher | View at Google Scholar - Carvell MJ, Persaud SJ, Jones PM. An islet is greater than the sum of its parts: the importance of intercellular communication in insulin secretion. Cell Science Rev 2006 ; 3: 100 – 128.
View at Publisher | View at Google Scholar - Ravier MA , Guldenagel M , Charollais A , Gjinovci A , Caille D , Sohl G, et al. Loss of connexin 36 channels alters beta-cell coupling, islet synchronization of glucose-induced Ca 2+ and insulin oscillations, and basal insulin release. Diabetes 2005; 54: 1798 – 1807.
View at Publisher | View at Google Scholar - Serre - Beinier V , Bosco D , Zulianello L , Charollais A , Caille D , Charpantier E , et al. Cx36 makes channels coupling human pancreatic beta-cells and correlates with insulin expression. Hum Mol Genet 2009 ; 18 : 428 – 439
View at Publisher | View at Google Scholar - Carvell MJ, Marsh PJ, Persaud SJ, Jones PM. E - E-cadherin interactions alter beta-cellular proliferation in islet-like systems. mobile Physiol Biochem 2007; 20: 617 – 626.
View at Publisher | View at Google Scholar - Jaques F, Jousset H, Tomas A, Prost AL, Wollheim CB, Irminger JC, et al. Dual effect of mobile–mobile contact disruption on cytosolic calcium and insulin secretion. Endocrinology 2008; 149: 2494 – 2505.
View at Publisher | View at Google Scholar - Konstantinova I , Nikolova G , Ohara - Imaizumi M , Meda P , Kucera T , Zarbalis ok , et al. EphA - Ephrins - A - mediated beta cellular communique regulates insulin secretion from pancreatic islets. mobile 2007; 129: 359 – 370.
View at Publisher | View at Google Scholar - Van Schravendijk CF , Foriers A , Hooghe - Peters EL , Rogiers V , De MP , Sodoyez JC , et al. Pancreatic hormone receptors on islet cells. Endocrinology 1985; 117: 841 – 848.
View at Publisher | View at Google Scholar - Hauge - Evans AC, King AJ, Carmignac D, Richardson CC, Robinson IC, Low MJ, et al. Somatostatin secreted by using islet delta-cells fulfills more than one role as a paracrine regulator of the islet feature. Diabetes 2009; 58: 403 – 411.
View at Publisher | View at Google Scholar - Ravier MA , Rutter GA . Glucose or insulin, however not zinc ions, inhibit glucagon secretion from mouse pancreatic alpha-cells. Diabetes 2005; 54: 1789 – 1797.
View at Publisher | View at Google Scholar - Persaud SJ , Muller D , Jones PM . Insulin signaling in islets. Biochem Soc Trans 2008; 36: 290 – 293.
View at Publisher | View at Google Scholar - Kitsou - Mylona I, Burns CJ, Squires PE, Persaud SJ, Jones PM. A function for the extracellular calcium-sensing receptor in cellular–cellular verbal exchange in pancreatic islets of Langerhans. mobile Physiol Biochem 2008; 22: 557 – 566.
View at Publisher | View at Google Scholar - Detimary P, Jonas JC, Henquin JC. stable and diffusible pools of nucleotides in pancreatic islet cells. Endocrinology 1996; 137: 4671 – 4676.
View at Publisher | View at Google Scholar - Rorsman P , Berggren PO , Bokvist ok , Ericson H , Mohler H , Ostenson CG, et al. Glucose - inhibition of glucagon secretion involves activation of GABAA - receptor chloride channels. Nature 1989; 341: 233 –236
View at Publisher | View at Google Scholar - Rubi B, Ljubicic S, Pournourmohammadi S, Carobbio S, Armanet M, Bartley C, et al. Dopamine D2-like receptors are expressed in pancreatic beta cells and mediate the inhibition of insulin secretion. J Biol Chem 2005; 280: 36824 – 36832.
View at Publisher | View at Google Scholar - Hellman B, Grapengiesser E, Gylfe E . Nitric oxide: a putative syn synchronizer of pancreatic beta-cell activity. Diabetes Res Clin Pract 2000; 50: S148.
View at Publisher | View at Google Scholar - Henningsson R, Alm P, Ekstrom P, Lundquist I . Heme oxygenase and carbon monoxide: regulatory roles in islet hormone release – a biochemical, immuno histochemical, and confocal microscopic study. Diabetes 1999; 48: 66 – 76.
View at Publisher | View at Google Scholar - Ramracheya RD, Muller DS, Wu Y, Whitehouse BJ, Huang GC, Amiel SA, et al. Direct regulation of insulin secretion by angiotensin II in human islets of Langerhans. Diabetologia 2006 ; 49 : 321 – 331 .
View at Publisher | View at Google Scholar - Hauge - Evans AC, Richardson CC, Milne HM, Christie MR, Persaud SJ, Jones PM. A role for kisspeptin in islet function. Diabetologia 2006; 49: 2131 – 2135.
View at Publisher | View at Google Scholar - Bell GI , Pictet RL , Rutter WJ , Cordell B , Tischer E , Goodman HM . The sequence of the human insulin gene. Nature 1980; 284: 26 – 32.
View at Publisher | View at Google Scholar - Hutton JC. Insulin secretory granule biogenesis and the proinsulin-processing endopeptidases. Diabetologia 1994 ; 37 ( Suppl 2 ): S48 – S56 .
View at Publisher | View at Google Scholar - Andrali SS, Sampley ML, Vanderford NL, Ozcan S . Glucose regulation of insulin gene expression in pancreatic beta-cells. Biochem J 2008; 415: 1 – 10.
View at Publisher | View at Google Scholar - Ashcroft SJ, Bunce J, Lowry M, Hansen SE, Hedeskov CJ. The effect of sugars on (pro)insulin biosynthesis. Biochem J 1978; 174: 517 – 526.
View at Publisher | View at Google Scholar - Hutton JC. The insulin secretory granule. Diabetologia 1989 ; 32 : 271 – 281.
View at Publisher | View at Google Scholar - Hickey AJ, Bradley JW, Skea GL, Middle ditch MJ, Buchanan CM, Phillips AR, et al. Proteins associated with immuno purified granules from a model pancreatic islet beta-cell system: a proteomic snapshot of an endocrine secretory granule. J Proteome Res 2009; 8: 178 – 186.
View at Publisher | View at Google Scholar - Howell SL, Young DA, Lacy PE. Isolation and properties of secretory granules from rat islets of Langerhans. 3. Studies of the stability of the isolated beta granules. J Cell Biol 1969; 41: 167 – 176.
View at Publisher | View at Google Scholar - Dunn MF. Zinc – ligand interactions modulate assembly and stability of the insulin hexamer: a review. Bio metals 2005; 18: 295 – 303.
View at Publisher | View at Google Scholar - Li G , Rungger - Brandle E , Just I , Jonas JC , Aktories K , Wollheim CB . Effect of disruption of actin fi laments by Clostridium botulinum C2 toxin on insulin secretion in HIT - T15 cells and pancreatic islets. Mol Biol Cell 1994; 5: 1199 – 1213.
View at Publisher | View at Google Scholar - Thurmond DC , Gonelle - Gispert C , Furukawa M , Halban PA , Pessin JE. Glucose-stimulated insulin secretion is coupled to the interaction of actin with the t - SNARE (target membrane soluble N - ethyl maleimide - sensitive factor attachment protein receptor protein) complex. Mol Endocrinol 2003; 17: 732 – 742.
View at Publisher | View at Google Scholar - Boyd AE III, Bolton WE, Brinkley BR. Microtubules and beta cell function: effect of colchicine on microtubules and insulin secretion in vitro by mouse beta cells. J Cell Biol 1982; 92: 425 – 434.
View at Publisher | View at Google Scholar - Howell SL, Tyhurst M. The cytoskeleton and insulin secretion. Diabetes Metab Rev 1986; 2: 107 – 123.
View at Publisher | View at Google Scholar - Howell SL. The mechanism of insulin secretion. Diabetologia 1984 ; 26: 319 – 327.
View at Publisher | View at Google Scholar - Meng YX, Wilson GW, Avery MC, Varden CH, Balczon R. Suppression of the expression of a pancreatic beta-cell form of the kinesin heavy chain by antisense oligonucleotides inhibits insulin secretion from primary cultures of mouse beta-cells. Endocrinology 1997; 138: 1979 – 1987.
View at Publisher | View at Google Scholar - Varadi A , Ainscow EK , Allan VJ , Rutter GA . Molecular mechanisms involved in secretory vesicle recruitment to the plasma membrane in beta-cells. Biochem Soc Trans 2002; 30: 328 – 332.
View at Publisher | View at Google Scholar - Bai JZ , Mon Y , Krissansen GW . Kinectin participates in microtubule-dependent hormone secretion in pancreatic islet beta-cells. Cell Biol Int 2006; 30: 885 – 894.
View at Publisher | View at Google Scholar - Gauthier BR , Wollheim CB . Synaptotagmins bind calcium to release insulin. Am J Physiol Endocrinol Metab 2008; 295: E1279 – E1286.
View at Publisher | View at Google Scholar - Eliasson L, Abdulkader F, Braun M, Galvanovskis J, Hoppa MB, Rorsman P . Novel aspects of the molecular mechanisms controlling insulin secretion. J Physiol 2008; 586: 3313 – 3324.
View at Publisher | View at Google Scholar - Gut A, Kiraly CE, Fukuda M, Mikoshiba K, Wollheim CB, Lang J . Expression and localization of synaptotagmin isoforms in endocrine beta-cells: their function in insulin exocytosis. J Cell Sci 2001 ; 114: 1709 – 1716.
View at Publisher | View at Google Scholar - De VA , Heimberg H , Quartier E , Huypens P , Bouwens L , Pipeleers D, et al. Human and rat beta cells differ in glucose transporter but not in glucokinase gene expression. J Clin Invest 1995; 96: 2489 – 2495.
View at Publisher | View at Google Scholar - Richardson CC, Hussain K, Jones PM, Persaud S, Lobner K, Boehm A, et al. Low levels of glucose transporters and K + ATP channels in human pancreatic beta cells early in improvement. Diabetologia 2007; 50: 1000 – 1005.
View at Publisher | View at Google Scholar - Van SE. quick-period regulation of glucokinase. Diabetologia 1994 ; 37 ( Suppl 2 ): S43 – S47 .
View at Publisher | View at Google Scholar - prepare dinner DL, Hales CN. Intracellular ATP straight away blocks k +channels in pancreatic B - B-cells. Nature 1984; 311: 271 – 273.
View at Publisher | View at Google Scholar - Miki T, Nagashima K, Seino S . The shape and functions of the ATP - touchy okay + channel in insulin-secreting pancreatic beta-cells. J Mol Endocrinol 1999 ; 22 : 113 – 123 .
View at Publisher | View at Google Scholar - Zhang Y, Warren-Perry M, Sakura H, Adelman J, Stoffel M, Bell GI, et al. No proof for mutations in a putative beta - mobile ATP - sensitive okay + channel subunit in MODY, NIDDM, or GDM. Diabetes 1995; 44: 597 – 600.
View at Publisher | View at Google Scholar - Sakura H, Wat N, Horton V, Millns H, Turner RC, Ashcroft FM. series versions in the human Kir6.2 gene, a subunit of the beta-cell ATP - touchy ok - channel – no association with NIDDM in a while Caucasian subjects or evidence of bizarre characteristics even as expressed in vitro. Diabetologia 1996 ; 39 : 1233 – 1236 .
View at Publisher | View at Google Scholar - Gloyn AL, Weedon MN, Owen KR, Turner MJ, Knight BA, Hitman G, et al. Large-scale association research of variants in genes encoding the pancreatic beta-cellular KATP channel subunits Kir6.2 (KCNJ11) and SUR1 (ABCC8) confirm that the KCNJ11 E23K variation is associated with type 2 diabetes. Diabetes 2003; fifty-two: 568 – 572.
View at Publisher | View at Google Scholar - Sagen JV, Raeder H, Hathout E, Shehadeh N, Gudmundsson K, Baevre H, et al. Everlasting neonatal diabetes because of mutations in KCNJ11 encoding Kir6.2: patient trends and preliminary reaction to sulfonylurea therapy. Diabetes 2004; -53: 2713 – 2718.
View at Publisher | View at Google Scholar - Thomas PM, Cote GJ, Wohllk N, Haddad B, Mathew PM, Rabl W, et al. Mutations within the sulfonylurea receptor gene in familial continual Hyperinsulinemic hypoglycemia of infancy. technology 1995; 268: 426 – 429.
View at Publisher | View at Google Scholar - Sharma N, Crane A, Gonzalez G, Bryan J, Guilar - Bryan L . Familial hyperinsulinism and pancreatic beta - cellular ATP - sensitive potassium channels. Kidney Int 2000; fifty-seven: 803 – 808.
View at Publisher | View at Google Scholar - Barg S, Ma X, Eliasson L, Galvanovskis J, Gopel SO, Obermuller S, et al. Fast exocytosis with few Ca 2+ channels in insulin-secreting mouse pancreatic B cells. Biophys J 2001; eighty-one: 3308 – 3323.
View at Publisher | View at Google Scholar - Jones PM, Persaud SJ, Howell SL. Time - direction of Ca 2+ - brought on insulin secretion from perifused, electrically permeabilized islets of Langerhans: consequences of cAMP and a phorbol ester. Biochem Biophys Res Commun 1989; 162: 998 – 1003.
View at Publisher | View at Google Scholar - Thore S, Wuttke A, Tengholm A . Fast turnover of phosphatidylinositol -4, 5 - bisphosphate in insulin-secreting cells mediated through Ca 2+and the ATP - to - ADP ratio. Diabetes 2007; 56: 818 – 826.
View at Publisher | View at Google Scholar - Jones PM , Persaud SJ . Protein kinases, protein phosphorylation, and the regulation of insulin secretion from pancreatic beta-cells. Endocr Rev 1998; 19: 429 – 461.
View at Publisher | View at Google Scholar - Robertson RP. The dominance of cyclooxygenase - 2 in the law of pancreatic islet prostaglandin synthesis. Diabetes 1998; 47: 1379 – 1383.
View at Publisher | View at Google Scholar - Persaud SJ, Muller D, Belin VD, Papadimitriou A, Huang GC, Amiel SA, et al. Expression and function of cyclooxygenase and lipoxygenase enzymes in human islets of Langerhans. Arch Physiol Biochem 2007; 113: 104 – 109.
View at Publisher | View at Google Scholar - Tian YM, Urquidi V, Ashcroft SJ. Protein kinase C in beta-cells: expression of a couple of isoforms and involvement in cholinergic stimulation of insulin secretion. Mol Cell Endocrinol 1996 ; 119 : 185 –193
View at Publisher | View at Google Scholar - Biden TJ , Schmitz - Peiffer C , Burchfi eld JG , Gurisik E , Cantley J , Mitchell CJ, et al. The various roles of protein kinase C in pancreatic beta-cell function. Biochem Soc Trans 2008; 36: 916 – 919.
View at Publisher | View at Google Scholar - Szaszak M, Christian F, Rosenthal W, Klussmann E . Compartmentalized cAMP signaling in regulated exocytic strategies in non-neuronal cells. mobile sign 2008; 20: 590 – 601.
View at Publisher | View at Google Scholar - Persaud SJ, Jones PM, Howell SL. Glucose-inspired insulin secretion isn't always dependent on the activation of protein kinase A . Biochem Biophys Res Commun 1990; 173: 833 – 839.
View at Publisher | View at Google Scholar - Dyachok O, Idevall - Hagren O, Sagetorp J, Tian G, Wuttke A, Arrieumerlou C , et al. Glucose-brought about cyclic AMP oscillations adjust pulsatile insulin secretion. mobile Metab 2008; 8: 26 – 37.
View at Publisher | View at Google Scholar - Henquin JC. Triggering and amplifying pathways of regulation of insulin secretion by
View at Publisher | View at Google Scholar - Gembal M , Gilon P , Henquin JC . evidence that glucose can manage insulin release independently from its motion on ATP - ATP-touchy K channels in mouse B cells. J Clin makes investments 1992; 89: 1288 – 1295.
View at Publisher | View at Google Scholar - Ravier MA , Nenquin M , Miki T , Seino S , Henquin JC . Glucose controls cytosolic Ca 2+ and insulin secretion in mouse islets lacking adenosine triphosphate-touchy k+ +channels owing to a knockout of the pore-forming subunit Kir6.2. Endocrinology 2009; 150: 33 – 45.
View at Publisher | View at Google Scholar - Detimary P , Gilon P , Nenquin M , Henquin JC . sites of glucose manipulation of insulin launch with wonderful dependence on the powerful nation in pancreatic B - B-cells. Biochem J 1994; 297 (part three): 455 – 461.
View at Publisher | View at Google Scholar - Henquin JC , Meissner HP . Cyclic adenosine monophosphate differently impacts the response of mouse pancreatic beta-cells to numerous amino acids. J Physiol 1986; 381: 77 – 93.
View at Publisher | View at Google Scholar - Minami ok, Miki T, Kadowaki T, Seino S . Roles of ATP - sensitive okay +channels as metabolic sensors: studies of Kir6.x null mice. Diabetes 2004 ; 53 ( Suppl 3 ): S176 – S180 .
View at Publisher | View at Google Scholar - Muller D, Huang GC, Amiel S, Jones PM, Persaud SJ. identity of insulin signaling factors in human beta-cells: autocrine law of insulin gene expression. Diabetes 2006; 55 2835 – 2842.
View at Publisher | View at Google Scholar - Khan FA, Goforth PB, Zhang M, Satin LS. Insulin activates ATP - touchy k + channels in pancreatic beta-cells through a phosphatidylinositol three - kinase-established pathway. Diabetes 2001; 50: 2192 – 2198.
View at Publisher | View at Google Scholar - Zawalich WS , Zawalich KC . effects of glucose, exogenous insulin, and carbachol on C - peptide and insulin secretion from remoted perfused rat islets. J Biol Chem 2002; 277: 26233 – 26237.
View at Publisher | View at Google Scholar - Leibiger B , Leibiger IB , Moede T , Kemper S , Kulkarni RN , Kahn CR , et al. Selective insulin signaling via A and B insulin receptors regulates the transcription of insulin and glucokinase genes in pancreatic beta cells. Mol mobile 2001; 7: 559 – 570.
View at Publisher | View at Google Scholar - Muller D , Jones PM , Persaud SJ . Autocrine anti-apoptotic and proliferative outcomes of insulin in pancreatic beta-cells. FEBS Lett 2006; 580: 6977 – 6980.
View at Publisher | View at Google Scholar - Gromada J , Franklin I , Wollheim CB . Alpha-cells of the endocrine pancreas: 35 years of research but the enigma remains. Endocr Rev 2007; 28: 84 – 116.
View at Publisher | View at Google Scholar - Williams G, Bloom SR. Somatostatin and pancreatic polypeptide. In: Albert KGMM, DeFronzo RA, keen H, Zimmett P, eds. International Textbook of Diabetes Mellitus. Chichester: Wiley, 1992; 341 – 355.
View at Publisher | View at Google Scholar - Holst JJ, Orskov C, Bersani M . Heterogeneity of islet hormones. In: Samols E, ed. The Endocrine Pancreas. Big Apple: Raven Press, 1991; 125 – 152.
View at Publisher | View at Google Scholar - Vieira E, Salehi A, Gylfe E . Glucose inhibits glucagon secretion using a direct impact on mouse pancreatic alpha cells. Diabetologia 2007 ; 50: 370 – 379.
View at Publisher | View at Google Scholar - Kumar U , Sasi R , Suresh S , Patel A , Thangaraju M , Metrakos P , et al. Subtype - a selective expression of the five somatostatin receptors (hSSTR1 - 5) in human pancreatic islet cells: a quantitative double-label immuno histochemical analysis. Diabetes 1999; 48: 77 – 85
View at Publisher | View at Google Scholar - Strowski MZ, Parmar RM, Blake ad, Schaeffer JM. Somatostatin inhibits insulin and glucagon secretion via receptors subtypes: an in vitro examination of pancreatic islets from somatostatin receptor 2 knockout mice. Endocrinology 2000; 141: 111 –117.
View at Publisher | View at Google Scholar - Sharp GW. Mechanisms of inhibition of insulin launch. Am J Physiol 1996; 271: C1781 – C1799.
View at Publisher | View at Google Scholar - Nilsson T , Arkhammar P , Rorsman P , Berggren PO . Suppression of insulin launch by galanin and somatostatin is mediated by using a G - protein: an effect concerning repolarization and reduction in cytoplasmic unfastened Ca 2+ attention. J Biol Chem 1989; 264: 973 – 980.
View at Publisher | View at Google Scholar - Degano P, Peiro E, Miralles P, Silvestre RA, Marco J . Outcomes of rat pancreatic polypeptide on islet-cell secretion within the perfused rat pancreas. Metabolism 1992; 41: 306 – 309.
View at Publisher | View at Google Scholar - Wierup N , Svensson H , Mulder H , Sundler F . The Ghrelin mobile: a novel developmentally regulated islet mobile inside the human pancreas. Regular Pept 2002; 107: 63 – 69.
View at Publisher | View at Google Scholar - Andralojc KM , Mercalli A , Nowak KW , Albarello L , Calcagno R , Luzi L, et al. Ghrelin-generating epsilon cells within the developing and grown human pancreas. Diabetologia 2009; fifty-two: 486 – 493.
View at Publisher | View at Google Scholar - Dezaki ok, Sone H, Yada T . Ghrelin is a physiological regulator of insulin launch in pancreatic islets and glucose homeostasis. Pharmacol Ther 2008; 118: 239 – 249.
View at Publisher | View at Google Scholar - Gentes M . notice sur les terminaisons nerveuses des islots de Langerhans du pancreas. Soc Biol 1902; 54: 202 – 203.
View at Publisher | View at Google Scholar - Gilon P , Henquin JC . Mechanisms and physiological significance of the cholinergic manipulation of pancreatic beta-cell function. Endocr Rev 2001; 22: 565 – 604.
View at Publisher | View at Google Scholar - Porte D Jr, Seeley RJ, Woods SC, Baskin DG, Figlewicz DP, Schwartz MW .weight problems, diabetes, and the significant fearful gadget. Diabetologia1998; 41: 863 – 881.
View at Publisher | View at Google Scholar - Iismaa TP, Kerr EA, Wilson JR, woodworker L, Sims N, Biden TJ. Quantitative and purposeful characterization of muscarinic receptor subtypes in insulin-secreting cellular strains and rat pancreatic islets. Diabetes 2000; 49: 392 – 398.
View at Publisher | View at Google Scholar - Ahren B, Lundquist I. Results of selective and non-selective beta-adrenergic retailers on insulin secretion in vivo. Eur J Pharmacol 1981 ; 71: 93 – 104.
View at Publisher | View at Google Scholar - Morgan NG, Chan SL, Lacey RJ, Brown CA. Pharmacology and molecular biology of islet cellular adrenoceptor. In: Flatt PR , Lenzen S , eds. Frontiers of Insulin Secretion and Pancreatic B - cell research. London: Smith - Gordon, 1994; 359 – 368.
View at Publisher | View at Google Scholar - Lacey RJ, Berrow NS, London NJ, Lake SP, James RF, Scarpello JH, et al. Differential effects of beta-adrenergic agonists on insulin secretion and pancreatic islets isolated from rats and guys. J Mol Endocrinol 1990 ; 5 : 49 – 54 .
View at Publisher | View at Google Scholar - Kuo WN, Hodgins DS, Kuo JF. Adenylate cyclase in islets of Langerhans: isolation of islets and the law of adenylate cyclase hobby by using various hormones and retailers. J Biol Chem 1973; 248: 2705 – 2711.
View at Publisher | View at Google Scholar - Yamazaki S, Katada T, Ui M . Alpha 2 - adrenergic inhibition of insulin secretion through interference with cyclic AMP era in rat pancreatic islets. Mol Pharmacol 1982 ; 21 : 648 – 653 .
View at Publisher | View at Google Scholar - Jones PM, Fyles JM, Persaud SJ, Howell SL. Catecholamine inhibition of Ca 2+ - brought on insulin secretion from electrically permeabilized islets of Langerhans. FEBS Lett 1987; 219: 139 – 144.
View at Publisher | View at Google Scholar - Winzell MS, Ahren B . G - protein-coupled receptors and islet function - implications for treatment of kind 2 diabetes. Pharmacol Ther 2007; 116: 437 – 448.
View at Publisher | View at Google Scholar - Wahl MA , Straub SG , Ammon HP . Vasoactive intestinal polypeptide - augmented insulin release: movements on ionic fluxes and electrical activity of mouse islets. Diabetologia 1993 ; 36 : 920 – 925 .
View at Publisher | View at Google Scholar - Straub SG , Sharp GW . Mechanisms of movement of VIP and PACAP in the stimulation of insulin release. Ann N Y Acad Sci 1996; 805: 607 – 612.
View at Publisher | View at Google Scholar - Wahl MA , Plehn RJ , Landsbeck EA , Verspohl EJ , Ammon HP .Gastrin-releasing peptide augments glucose-mediated 45Ca 2+ uptake, electric pastime, and insulin secretion of mouse pancreatic islets. Endocrinology 1991; 128: 3247 – 3252.
View at Publisher | View at Google Scholar - Dunning BE, Ahren B, Veith RC, Bottcher G, Sundler F, Taborsky GJ Jr . Galanin: a novel pancreatic neuro-peptide. Am J Physiol 1986; 251: E127 – E133.
View at Publisher | View at Google Scholar - Moltz JH , McDonald JK . Neuropeptides Y: direct and indirect motion on insulin secretion inside the rat. Peptides 1985; 6: 1155 – 1159.
View at Publisher | View at Google Scholar - Morgan DG, Kulkarni RN, Hurley JD, Wang ZL, Wang RM, Ghatei MA, et al. Inhibition of glucose-stimulated insulin secretion using neuro peptide Y is mediated via the Y1 receptor and inhibition of adenylyl cyclase in RIN 5AH rat insulinoma cells. Diabetologia 1998 ; 41: 1482 – 1491.
View at Publisher | View at Google Scholar - Cormont M, Le Marchand - Brustel Y, van OE, Spiegel AM, Sharp GW. Identification of G protein alpha-subunits in RINm5F cells and their selective interaction with galanin receptor. Diabetes 1991; 40: 1170 – 1176.
View at Publisher | View at Google Scholar - McIntyre N, Holds worth CD, Turner DS. A new interpretation of oral glucose tolerance. Lancet 1964; 2: 20 – 21.
View at Publisher | View at Google Scholar - Drucker DJ. The function of gut hormones in glucose homeostasis. J Clin invest 2007; 117: 24 – 32.
View at Publisher | View at Google Scholar - Karhunen LJ , Juvonen KR , Huotari A , Purhonen AK , Herzig KH . impact of protein, fats, carbohydrates, and fiber on gastrointestinal peptide release in people. Regular Pept 2008; 149: 70 – 78.
View at Publisher | View at Google Scholar - Kieffer TJ , Habener JF . The glucagon-like peptides. Endocr Rev 1999; 20: 876 – 913.
View at Publisher | View at Google Scholar - Drucker DJ , Nauck MA . The incretin machine: glucagon-like peptide - 1 receptor agonists and dipeptidyl peptidase - four inhibitors in type 2 diabetes. Lancet 2006; 368: 1696 – 1705.
View at Publisher | View at Google Scholar - Kolterman OG, Kim DD, Shen L, Ruggles JA, Nielsen LL, Fineman MS, et al. Pharmacokinetics, pharmacodynamics, and safety of exenatide in sufferers with type 2 diabetes mellitus. Am J Fitness Syst Pharm 2005; 62: 173 – 181.
View at Publisher | View at Google Scholar - Suga S, Kanno T, Ogawa Y, Takeo T, Kamimura N, Wakui M . cAMP - an impartial decrease of ATP - touchy okay + channel activity using GLP - 1 in rat pancreatic beta-cells. Pfl ugers Arch 2000 ; 440 : 566 – 572.
View at Publisher | View at Google Scholar - Ehses JA, Lee SS, Pederson RA, McIntosh CH. A brand new pathway for glucose-dependent insulinotropic polypeptide (GIP) receptor signaling: proof for the involvement of phospholipase A2 in GIP-inspired insulin secretion. J Biol Chem 2001; 276: 23667 – 23673.
View at Publisher | View at Google Scholar - Chia CW, Carlson OD, Kim W, Shin YK, Charles CP, Kim HS, et al. Exogenous glucose-structured insulinotropic polypeptide worsens postprandial hyperglycemia in type 2 diabetes. Diabetes 2009; 58: 1342 – 1349.
View at Publisher | View at Google Scholar - Morgan LM. The entire - insular axis. Biochem Soc Trans 1980; 8: 17 – 19.
View at Publisher | View at Google Scholar - Persaud SJ, Jones PM, Howell SL. Stimulation of insulin secretion by cholecystokinin - 8S: the position of protein kinase C . Pharmacol Commun 1993; 3: 39 – 47.
View at Publisher | View at Google Scholar - Catalan V , Gomez - Ambrosi J , Rodriguez A , Salvador J , Fruhbeck G . Adipokines within the treatment of diabetes mellitus and weight problems. professional Opin Pharmacother 2009; 10: 239 – 254.
View at Publisher | View at Google Scholar - instances JA , Gabriely I , Ma XH , Yang XM , Michaeli T , Fleischer N , et al. The physiological boom in plasma leptin markedly inhibits insulin secretion in vivo. Diabetes 2001; 50: 348 – 352.
View at Publisher | View at Google Scholar - Morioka T , Asilmaz E , Hu J , Dishinger JF , Kurpad AJ , Elias CF , et al. Disruption of leptin receptor expression inside the pancreas directly impacts beta-mobile growth and function in mice. J Clin Invest 2007; 117: 2860 – 2868.
View at Publisher | View at Google Scholar - Kieffer TJ , Heller RS , Leech CA , Holz GG , Habener JF . Leptin suppression of insulin secretion by using the activation of ATP-sensitive ok channels in pancreatic beta-cells. Diabetes 1997; 46: 1087 – 1093.
View at Publisher | View at Google Scholar - Maedler okay , Schulthess feet , Bielman C , Berney T , Bonny C , Prentki M, et al. Glucose and leptin set off apoptosis in human beta-cells and impaired glucose-stimulated insulin secretion via activation of c - Jun N - terminal kinases. FASEB J 2008 ; 22 : 1905 – 1913 .
View at Publisher | View at Google Scholar - Nakata M, Okada T, Ozawa K, Yada T . Resistin induces insulin resistance in pancreatic islets to impair glucose-induced insulin launch. Biochem Biophys Res Commun 2007; 353: 1046 – 1051.
View at Publisher | View at Google Scholar - Gao CL, Zhao DY, Qiu J, Zhang CM, Ji CB, Chen XH, et al. Resistin induces rat insulinoma cell RINm5F apoptosis. Mol Biol Rep 2009; 36: 1703 – 1708.
View at Publisher | View at Google Scholar - Arner P . Resistin: yet another adipokine tells us that men are not mice. Diabetologia 2005 ; 48 : 2203 – 2205 .
View at Publisher | View at Google Scholar - Minn AH , Patterson NB , Pack S , Hoffmann SC , Gavrilova O , Vinson C, et al. Resistin is expressed in pancreatic islets. Biochem Biophys Res Commun 2003; 310: 641 – 645.
View at Publisher | View at Google Scholar - Hotta K , Funahashi T , Arita Y , Takahashi M , Matsuda M , Okamoto Y, et al. Plasma concentrations of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc Biol 2000; 20: 1595 – 1599.
View at Publisher | View at Google Scholar - Gu W , Li X , Liu C , Yang J , Ye L , Tang J , et al. Globular adiponectin augments insulin secretion from pancreatic islet beta cells at high glucose concentrations. Endocrine 2006; 30: 217 – 221.
View at Publisher | View at Google Scholar - Okamoto M , Ohara - Imaizumi M , Kubota N , Hashimoto S , Eto K , Kanno T, et al. Adiponectin induces insulin secretion in vitro and in vivo at a low glucose concentration. Diabetologia 2008 ; 51: 827 – 835.
View at Publisher | View at Google Scholar - Rakatzi I, Mueller H, Ritzeler O, Tennagels N, Eckel J. (2004). Adiponectin counteracts cytokine - and fatty acid-induced apoptosis in the pancreatic beta-cell line INS - 1. Diabetologia; 47: 249 – 258.
View at Publisher | View at Google Scholar
