

Research Article | DOI: https://doi.org/10.31579/2835-2882/035
Investigation of the Dark Beetle and Its Effect on The Removal of Fungi
1 Department of Biology, Faculty of Basic Sciences, Payam Noor University, Mashhad, Iran
2 Department of Biology, Faculty of Basic Sciences; Payam Noor University, Tehran, Iran
*Corresponding Author: Majid Rajabian; Department of Biology, Faculty of Basic Sciences; Payam Noor University, Tehran, Iran
Citation: Amir Mahmoudi, Majid Rajabian, (2023), Investigation of the Dark beetle and its effect on the removal of fungi, Clinical Research and Studies, 2(5); DOI:10.31579/2835-2882/035
Copyright: © 2023, Majid Rajabian. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 29 September 2023 | Accepted: 09 October 2023 | Published: 20 October 2023
Keywords: dark beetle; cereals; end glands; nutritional value; fungi; Tenbrio Molitor
Abstract
The capacity of a single genotype to exhibit variable phenotypes in response to changing environmental conditions (ie, phenotypic plasticity) is common in insects, see reviews of, for example, body size divergence, mating, and life history strategies, population dynamics, replacement and cessation morphologies are triggered by plastic responses to adverse factors such as low temperature or poor nutrition. Climatic ranges are generally associated with heterogeneous environments including microclimatic gradients. Species distributed over such diverse habitats have certain phenological and phenotypic plasticity to adapt without compromising their fitness or biological success. Ecological and evolutionary responses to abiotic change at environmental and temporal scales require the advancement of information. These will improve our capacity to predict how species will respond to local and global environmental change. Some black cockroaches emit an unpleasant odor through the end glands of the body, which is very effective in driving away predators, and this substance changes the color of their bodies. What does this substance include? It can have in its composition that this species repels predators and changes the color of its body, considering that this animal lives in a humid environment, the possibility of preventing the growth of fungi is reduced. And it stops to some extent.
Introduction
Dark beetles are dark-colored beetles that belong to a large group of insects in the Tenebrionidae family. This beetle has about 1500 different species, which are mostly found in temperate and tropical regions of the world, and most of them live in fields and forest areas. Dark beetles can reach 1 to 1.5 inches in length. The body of dark beetles is covered with a black, smooth, armor-like protective shell. Adult cockroaches have a pair of segmented antennae, slitted eyes, and three pairs of legs. Beetles are dark in color due to the fused wings (also known as elytra) that are sealed to the body. And under the title of flour worms, there are two types of black beetles, the yellow flour worm (Tenebrio Molitor) and the dark or small flour worm (Tenebrio obscurus), which are considered by the experts of the global food system to be potentially one of the most valuable food sources for humans. (Martin et al. 1976, Vantomme et al. 2012). Although mealybugs are considered a cereal pest in some countries, they have been used as the most common food and feed additives in poultry, livestock, and aquaculture production (Klasing
et al. 2000; Ballitoc and Sun 2013). ) recently investigated the population structure and phylogenetic position of T. Molitor using mitochondrial data (Liu and Wang) 2014; Zhang et al. 2016). While only one mitochondrial sequence of T. obscurus is available from GenBank (Bai et). (al. 2018) In the present study, we report the mitogenome of T. obscurus from the Zunyi family, which provides valuable information for molecular phylogenetics and population genetics.
Specimens of T. obscurus (JNU20160522) were prepared and maintained by the Cell Biology Laboratory of Jiangsu Normal University. The complete mitogenome of T. obscurus was amplified using a combination of universal primers (Zhang et al. 2016). and annotated using the MITOS web server. (Bernt et al. 2013) Phylogenetic trees were reconstructed from 26 Cucujiformia mitogenomes using Bayesian inference (BI) and maximum likelihood (ML) methods.

The complete mitogenome of T. obscurus (GenBank accession number: KY270783) is 15,509 bp in size, which is shorter than the T. obscurus mitogenome of the Guiyang population. It contains a set of 37 conserved mitochondrial genes including 13 protein-coding genes (PCGs22 tRNA), 2 rRNA genes, and an A + T-rich region. The gene arrangement is identical to the normal insect gene arrangement. The heavy chain encodes 23 genes, including 14 tRNAs and 9 PCGs, and the other 14 genes are encoded by the light chain. The general base composition of the genome is as follows: A 43.1% T, 29.4% C, 17.4%, G 10.2%, and with an A + T content of 72.4%. All PCGs start with the ATN codon. Four PCGs terminate with incomplete stop codons, while nine PCGs terminate with TAA and TAG. All traditional tRNA genes from 60 to 70 bp have cloverleaf secondary structure, except tRNASer (AGN). The size of 16S and 12S rRNA genes of T. obscurus is 1280 bp and 780 bp, respectively. A + T rich regions show distinct differences between Guiyang (1177bp) and Zunyi (902bp) populations,
which can be used as a powerful molecular marker for population investigation. Some dark-colored cockroaches produce a foul-smelling substance from the end glands of the body to repel predators. This substance changes the color of the body to brown (21,31,32,33).
Dark beetles feed on decaying plant or animal matter, although they also feed on live plants, sprouts, fruits, mushrooms, and grains. The type of organism of dark cockroaches is such that they are used in laboratories to study the ecology, ecology, genetics, and biology of insects. Dark beetles have a complete metamorphosis, including eggs, larvae, pupae, and adult insects, and the female dark beetle can have up to 500 eggs, which hatch after a few weeks. The larvae are golden brown and have a cylindrical body equipped with 6 legs, which are used to move on the ground (Figure 1) (31,32).

Figure 1: The external organ of the dark beetle
Dark-colored cockroaches live in fields and forest areas. Many species live in close proximity to humans, in parks, gardens, barns, and other man-made objects that provide sufficient food. Dark beetles are classified as pests of cereals and other stored products in most countries. Adult cockroaches feed on decaying plant and animal matter. They also eat live plants, sprouts, fruits, fungi, and grains.
These insects rarely drink water, because they can extract moisture from the food they eat. The natural enemies of black cockroaches are lizards, rodents, spiders, birds, and large cockroaches. Dark-colored beetles are common
model organisms in laboratories and classrooms studying insect ecology, genetics, and biology. They are large, have a short life cycle, and do not require much effort to maintain. It is also used as an ideal food source for laboratory animals such as insectivorous mammals (22).
For the maintenance of the larvae, the suitable environment for it includes a temperature of 25 degrees and a humidity between 50 and 60%. The larva sheds 9 to 20 times in two months. Then it retreats underground and enters the pupa (sleeping) stage. Two or three weeks later, the white adult insect emerges from the cocoon. After only one day, it reaches the mature black color (Figure 2) (10,11).
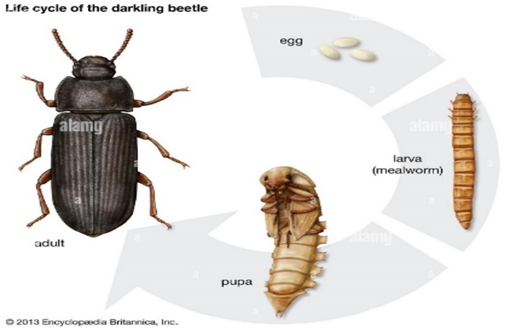
Figure 2: The life cycle of the Dark beetle
Some species of black cockroaches can complete their entire life cycle in 3 to 4 months, while others can live up to 10 years of age. Most dark-colored beetles live only a few months due to intense predation. Dark-colored beetles can be found under rocks, sand dunes, and deserts. Many are completely black, but some are auburn or brown. Some species eat mushrooms, others feed on live produce or decaying plant matter. Some are attracted to carrion, others are dung. Many are ignored and insignificant, but some are edible and produce products from them.
The range of behavior and life history among dark cockroaches is wide. Larvae are yellow-tan worm-like grubs. Commercially available food worm is the larva of a specific genus in the beetle family. Many species have a chemical defense as adults, they spray or secrete solutions from their bodies when they are anxious. Some cockroaches are long and narrow. Others have a banded belly or are round. While all have wings, species do not fly in arid regions because their wings are tightly packed, allowing them to prevent moisture from evaporating (Figure 3) (21).
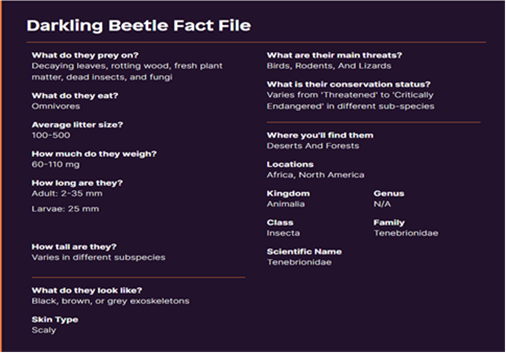
Figure 3: appearance and general information of the dark beetle
Research Methods
The nutritional value of the mealworm, Tenbrio Molitor was analyzed due to the increasing demand for its use as a protein source for pets and even more so for human consumption. The present work aimed to determine the chemical compost of Molitor Tenbrio larvae, adults, which were kept under standard conditions for further use of the mass rearing system and molting and its disposal. Tenebrio Molitor, larva, adult, molt, and its feces contained 46.44, 63.34, 32.87, and 18.51% of protein, respectively, suggesting that even excrement can be an additional supplement in the process of material recovery. Food to be used. This protein was also rich in amino acids such as isoleucine, leucine, and lysine, all of which met the nutritional value recommended by the Food and Agriculture Organization. Fatty acid composition with a high component of oleic acid, along with linoleic acid and palmyritic acid was detected in all adults, larvae, molts, and droppings. These components of oleic acid, linoleic acid, and palmyritic acid were the same or even strongly present in mealworm excrement of 22.29, 47.19, and 19.17%, respectively. Longer chains of unsaturated fatty acids consisting of two to three double bonds were recognized as the healthy product in large quantities. These results show new ways to consume food worms and their waste for animal and human consumption (1, 2).
However, with the rapid growth of the human population and the continued development of modern civilization, the environment is subject to destruction and the climate is getting warmer. Therefore, it is worth considering alternative food production, which requires a large energy input or Produces large amounts of waste products and greenhouse gases, which is Examples of animal products such as pigs, cows, or raising chickens. Growing insects for human food is less burdensome on the environment and cheaper, so the output compensates for the effort. Insect farming has many benefits. It is more efficient than raising large animals, requires much less space, and produces less waste. Also, insects have a better capacity to convert plants into animal biomass than large animals. Experts in many sciences are increasingly devoting more time to this issue. The Food and Agriculture Organization of the United Nations predicts that the human population will grow to 9 billion people by 2050, and these people will need a valuable source of food. The rapid growth of the human population in the second half of the 21st century may lead to food shortages, especially in animal proteins. In a meeting held by FAO in Rome in 2012, possible solutions to this problem were proposed, including the use of insects as food. It was also planned to hold the first international scientific conference on the topic of using insects as food. Scientists are working on developing insect processing technologies to obtain healthy food that can replace farm animal meat. The European Union has allocated a large budget and currently spends 3 million on studies on the nutritional value of insects (5, 6).
The nutritional components of food worms can be classified as "source" according to the thresholds of the World Health Organization and the Food and Agriculture Organization of the United Nations food labels (Figure 4). Worms have a high content of protein (13.68–22.32 g/100 g edible portion) and fat (8.90–19.94 g/100 g edible portion), as well as significant amounts of polyunsaturated fatty acids. provide the Worms are also classified as a source of zinc and are high in magnesium, but contain only low levels of calcium. In addition, mealworms can be considered a source of niacin and are high in pyridoxine, riboflavin, folate, and vitamin B12. The nutritional profile of mealworms compared to conventional meats revealed that mealworms have a significantly higher nutritional value than beef and chicken and are not significantly nutritionally balanced. They also provide a good source of all essential amino acids (17, 18, 19).
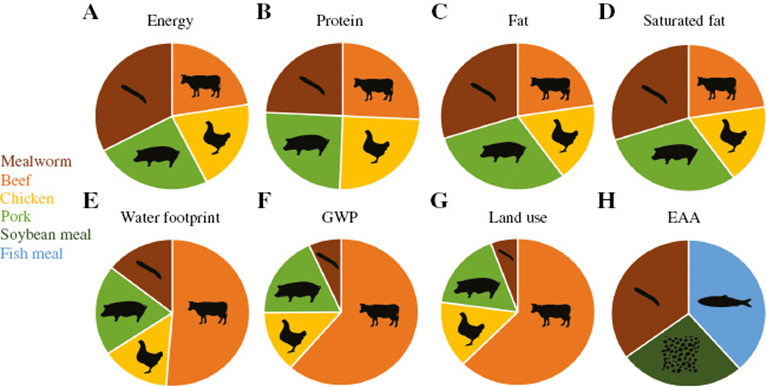
Figure 4: Nutritional values and stability of food worms and food/conventional food. (A-D) Nutritional value of a 100g edible portion of mealworms compared to different animal meats. (E–G) Comparative environmental impact of food worms and livestock presented as water footprint per ton of feed, global warming potential (GWP) per 1 kg feed portion, and land use per 1 kg Edible portion (H) essential amino acid (EAA) comparison of food worms and classic feeds such as Seva and fish meal.
Materials and methods
Since the dark beetle is used as a perfect source of protein for birds and other animals, we have thought that the gas released by this beetle is only used to drive away predators? The question arises that even though this cockroach can feed on mushrooms? Can it be used to kill stronger fungi and molds like Norspra and others like this type of mold? Does the proliferation of microorganisms decrease in the presence of the Dark beetle? What is the mechanism of action? By how many percent does it reduce the growth of microorganisms on "vegetable" food products?
To start the work, it is necessary to disinfect the equipment, and this is done with an autoclave. After disinfecting the equipment in a sterilized environment, we use agar plate count agar and mix 50 ml of 1.125 grams of agar in the bowl, after preparing the agar and contaminating it with the mold sample. We put the bread in the incubator for one day at a temperature of 25 degrees.
After a day has passed and the mold has grown in the petri dish, it is time to transfer it to the living environment of the dark cockroach. By placing the petri dish in the living environment and placing the cockroaches in the petri dish, we noticed that its thickness has decreased According to the evidence, the growth of mold in the Petri dish is completely stopped and no trace of mold can be seen.
To get more information that the applied substance was effective in stopping the growth of molds, we used the FT-IR (Fourier Transform Infrared) device (Figure 5). According to the analysis of the diagram, a part of this diagram is almost identical to the Agar diagram, but the only difference is in one peak, which despite the lack of facilities, there is the possibility of this theory that by dissecting this insect and detailed dissection We will come to a substance that can be used as a poison to repel insects and control the growth of molds (Figure 6).

Figure 5: Fourier Transform Infrared Spectroscopy – FTIR
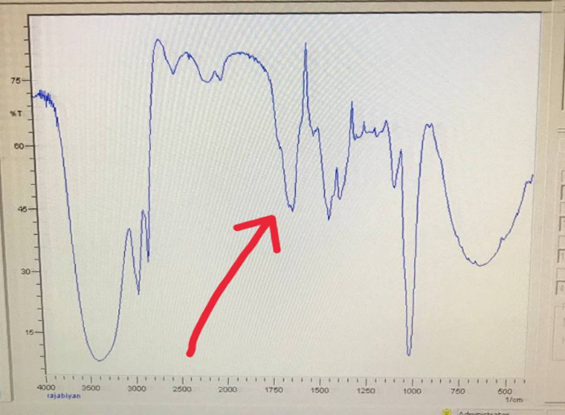
Figure 6: The result obtained from the FTIR device
According to this study, it represents the first identification of any diuretic agent from coleopteran insects. This cockroach can live in very dry conditions. Nicholson has suggested that in such beetles DH may act as a "cleansing hormone", providing fluid to the midgut to moisten dry food to aid digestion, the fluid being reabsorbed by the kryptonephridial (34).
However, we have named this peptide Tem-DHI, due to its effects on Mt and based on a nomenclature convention (35). The availability of synthetic Tem-DH1 would greatly aid physiological studies of diuresis in cockroaches. In this paper, a diuretic hormone with a structure was isolated from the extract of whole heads of the earthworm Tenebrio Molitor. This hormone is a 37-aa peptide of 4371 Da with the sequence SPTISITAPIDVLRKTWEQERARKQMVKNREFLNSLN.
This peptide increases cAMP production in Malpighian tubules of T. Molitor. The amino acid sequence indicates that this peptide is a member of the savagin/corticotropin/ortensin I-related family of insect urinary hormones. The C-terminal sequence of this peptide is completely different from other members of this family that have a hydrophobic C-terminus (isoleucinamide or valine amide). When the diuretic hormone of T. Molitor is similarly aligned, it has a more hydrophilic C terminus, lucilasparagine (free acid). Unlike other known diuretic hormones of this family, this peptide has exceptionally low stimulatory activity on cAMP production in Malpighian tubules of Manduca sexta. However, at nanomolar concentrations, it stimulates cAMP production in Malpighian tubules of T. Molitor. Diuretic hormones of this family have already been isolated from Lepidoptera, Orthoptera, Dictyoptera, and Diptera. It seems. This is the first diuretic hormone isolated from a coleopteran insect.
In a study conducted by Urba'nski and his colleagues (36), four types of hemocytes were observed: granulocytes, plasmatocytes, endocytosis, and prohemocytes according to their size and morphology. Prohemocytes are considered to stem cells capable of differentiating into other types of cells (12). In addition, granulocytes and plasmatocytes are the most abundant types of hemolymph. They are the only cells with adhesion properties, and the cells responsible for phagocytic activity (Figure 3), encapsulation, and modulation (37,38). The enzyme phenol oxidase (PO) plays a key role in mediating this important defense mechanism. This enzyme is the main component responsible for the process of melanization of hemocytes attached to the pathogen (39). Studies show that prophenol oxidase (Propo) is mainly synthesized by granulocyte and oenocytoid hemocytes. and it depends on the insect species. Cellular and humoral immunology of T. Molitor as a tool to study human immunity to protect against potentially pathogenic microbes, the skin, and gastrointestinal tract represents the first line of defense that plays an important role against infections. slow However, when these barriers are overcome, the invading pathogen is exposed to a wide range of cellular and humoral. Mechanisms (35). In a study by Urbański et al. (36), four types of hemocytes were observed: granulocytes, plasmatocytes, endocytosis, and prohemocytes according to size and morphology. Prohemocytes are considered stem cells capable of differentiating into other cell types (40). In addition, granulocytes and plasmatocytes are the most abundant types in hemolymph, the only cells with adhesive properties, and are the cells responsible for phagocytic activity (Figure 3), encapsulation, and modulation (37,38). The enzyme phenol oxidase (PO) plays a key role in mediating this important defense mechanism. This enzyme is the main component is responsible for the process of melanization of hemocytes attached to the pathogen (39). Studies show that prophenol oxidase (Propo) is pred.
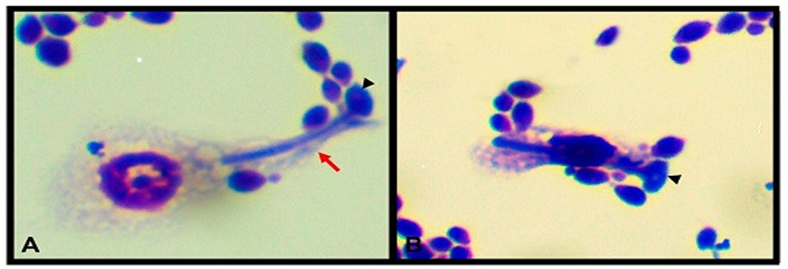
Figure 3. Candida albicans phagocytizing plasmatocyte (A) and granulocyte (B). Mi-Grunwaldgemsa staining and simultaneous incubation for 2 hours. Cytoplasmic protrusion (pseudo-red arrow) towards the mushroom (arrowhead) is observed. 100x objective lens.
References
- Aguilar-Miranda ED, Lopez MG, Escamilla-Santana C, Barba De La Rosa AP (2002).
View at Publisher | View at Google Scholar - Characteristics of maize flour tortilla supplemented with ground Tenebrio molitor larvae, J. Agric. Food Chem 50, 192-195.
View at Publisher | View at Google Scholar - Allen MO, Oftedal OT, Baer DJ (1996). The feeding and nutrition of carnivores; in wild mammals in captivity. Kleiman DG (ed), pp.139-147, Univ.of Chicago Press, Chicago, Illinois.
View at Publisher | View at Google Scholar - O. A. C (1990). Official Methods of Analysis (15th ed.). Association of official analytical chemists. Washington D C, USA.
View at Publisher | View at Google Scholar - O. A. C (2003). Official Methods of Analysis. Crude Fat in Feeds, Cereal Grains, and Forages. International USA.
View at Publisher | View at Google Scholar - Barker D, Fitzpatrick MP, Dierenfeld ES (1998). Nutrient compotion of selected whole invertebrates. Zoo Biology 17, 123-134.
View at Publisher | View at Google Scholar - Brues CT (1946). Insect Dietary. Harvard University Press, USA.
View at Publisher | View at Google Scholar - Bukkens SGF (1996). The nutritional value of edible insects. Ecology of Food and Nutrition 36, 287-319.
View at Publisher | View at Google Scholar - Capinera JL (2004). Cycloperdia of Entomology. Kluwer Academic press , Boston, USA.
View at Publisher | View at Google Scholar - Defoliart GR (2002). The Human Use of Insects as Food Resource; A Bibliographic Account in Progress. University of Wisconsin -Madison, Madison, Wis, USA.
View at Publisher | View at Google Scholar - Insects | Free Full-Text | Interpopulation Plasticity in a Darkling Beetle Life-History along a Whole Oceanic Island Altitudinal Gradient | HTML (mdpi.com)
View at Publisher | View at Google Scholar - Bark beetles: A natural and dramatic forest disturbance | Rocky Mountain Research Station (usda.gov)
View at Publisher | View at Google Scholar - Schiefenhövel, W. and Blum, P. (2009). Insects: Forgotten and rediscovered as ford. Entomophagy among the Eipo, highlands of West New Guinea, and in other traditional societies. In: MacClancy, J., Henry, J. and Macbeth, H., Eds., Consuming the Inedible, Berghaghn Books, New York, 163-176.
View at Publisher | View at Google Scholar - Johnson, D.V. (2010). The contribution of edible forest insects to human nutrition and forest management. In: Forest Insects as Food: Humans Bite Back, FAO of the United Nations Regional Office for Asia and the Pacific, Bangkok, 5-22FAO (2010) Forest insects as food: Humans bite back. FAO of the United Nations Regional Office for Asia and the Pacific, Bangkok.
View at Publisher | View at Google Scholar - Schiefenhövel, W. and Blum, P. (2009). Insects: Forgotten and rediscovered as ford. Entomophagy among the Eipo, highlands of West New Guinea, and in other traditional societies. In: MacClancy, J., Henry, J. and Macbeth, H., Eds., Consuming the Inedible, Berghaghn Books, New York, 163-176.
View at Publisher | View at Google Scholar - Johnson, D.V. (2010). The contribution of edible forest insects to human nutrition and forest management. In: Forest Insects as Food: Humans Bite Back, FAO of the United Nations Regional Office for Asia and the Pacific, Bangkok, 5-22FAO (2010) Forest insects as food: Humans bite back. FAO of the United Nations Regional Office for Asia and the Pacific, Bangkok.
View at Publisher | View at Google Scholar - Rumpold BA, Schlüter OK. (2013). Nutritional composition and safety aspects of edible insects. Mol Nutr Food Res
View at Publisher | View at Google Scholar - Nowak V, Persijn D, Rittenschober D, Charrondiere UR. (2016). Review of food composition data for edible insects. Food Chem 193:39–46.
View at Publisher | View at Google Scholar - Codex Alimentarius Commission (Ed.). (2007). Food Labelling 5th ed. World Health Organization [u.a.], Rome,
View at Publisher | View at Google Scholar - Payne CL, Scarborough P, Rayner M, Nonaka K. (2016). Are edible insects more or less “healthy” than commonly consumed meats? A comparison using two nutrient profiling models developed to combat over- and undernutrition. Eur J Clin Nutr
View at Publisher | View at Google Scholar - Hadley, Debbie.
View at Publisher | View at Google Scholar - Hadley, Debbie.
View at Publisher | View at Google Scholar - Bai Y, Li C, Yang M, Liang S. (2018). Complete mitochondrial genome of the dark mealworm Tenebrio obscurus Fabricius (Insecta: Coleoptera: Tenebrionidae). Mitochondrial DNA Part B. 3:171–172.
View at Publisher | View at Google Scholar - Ballitoc DA, Sun S. (2013). Ground yellow mealworms (Tenebrio molitor L.) feed supplementation improves growth performance and carcass yield characteristics in broilers. Open Sci Repo Agric. e23050425.
View at Publisher | View at Google Scholar - Bernt M, Donath A, Jühling F, Externbrink F, Florentz C et al. (2013). MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69:313–319.
View at Publisher | View at Google Scholar - Klasing KC, Thacker P, Lopez MA, Calvert CC. (2000). Increasing the calcium content of mealworms (Tenebrio molitor) to improve their nutritional value for bone mineralization of growing chicks. J Zoo Wildl Med. 31:512–517.
View at Publisher | View at Google Scholar - Liu LN, Wang CY. (2014). Complete mitochondrial genome of yellow meal worm (Tenebrio molitor). Zool Res. 35:537–545.
View at Publisher | View at Google Scholar - Martin RD, Rivers JPW, Cowgill UM. (1976). Culturing mealworms as food for animals in captivity. Int Zoo Yearb. 16:63–70.
View at Publisher | View at Google Scholar - Vantomme P, Mertens E, van Huis A, Klunder H. (2012). Assessing the potential of insects as food and feed in assuring food security: summary report. Rome (Italy): Food and Agricultural Organization of the United Nations, Forestry Department.
View at Publisher | View at Google Scholar - Zhang H, Liu N, Han Z, Liu J. (2016). Phylogenetic analyses and evolutionary timescale of Coleoptera based on mitochondrial sequence. Biochem Syst Ecol. 66:229–238.
View at Publisher | View at Google Scholar - Bai Y, Li C, Yang M, Liang S. (2018). Complete mitochondrial genome of the dark mealworm Tenebrio obscurus Fabricius (Insecta: Coleoptera: Tenebrionidae). Mitochondrial DNA Part B. 3:171–172.
View at Publisher | View at Google Scholar - Ballitoc DA, Sun S. (2013). Ground yellow mealworms (Tenebrio molitor L.) feed supplementation improves growth performance and carcass yield characteristics in broilers. Open Sci Repo Agric. e23050425.
View at Publisher | View at Google Scholar - Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, et al. (2013). MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69:313–319
View at Publisher | View at Google Scholar - Klasing KC, Thacker P, Lopez MA, Calvert CC. (2000). Increasing the calcium content of mealworms (Tenebrio molitor) to improve their nutritional value for bone mineralization of growing chicks. J Zoo Wildl Med. 31:512–517.
View at Publisher | View at Google Scholar - Tindwa, H.; Jo, Y.H.; Patnaik, B.B.; Lee, Y.S.; Kang, S.S.; et al. (2015). Molecular cloning and characterization of autophagy-related gene TmATG8 in Listeria-invaded hemocytes of Tenebrio molitor. Dev. Comp. Immunol. 51, 88–98.
View at Publisher | View at Google Scholar - . Philips, T.W.; Throne, J.E. (2010). Biorational Approaches to Managing Stored-Product Insect. Annu. Rev. Entomol. 55, 375–397.
View at Publisher | View at Google Scholar - References 1. Hendriksen, C.F. (2009). Replacement, reduction and refinement alternatives to animal use in vaccine potency measurement. Expert Rev. Vaccines 8, 313–322. Urba´nski, A.; Adamski, Z.; Rosi ´nski, G. Developmental changes in haemocyte morphology in response to Staphylococcus aureus and latex beads in the beetle Tenebrio molitor L. Micron 2018, 104, 8–20.
View at Publisher | View at Google Scholar - . Ratcliffe, N.A. (1993). Cellular defense responses of insects: Unresolved problems. In Parasites and Pathogens of Insects; Beckage, N.E., Thompson, S.N., Federici, B.A., Eds.; Academic Press: San Diego, CA, USA, Volume 1, pp. 267–304. ISBN 9780080916491.
View at Publisher | View at Google Scholar - Strand, M.R.; Pech, L.L. (1995). Immunological basis for compatibility in parasitoid–host relationships. Annu. Rev. Entomol. 40, 31–56.
View at Publisher | View at Google Scholar - 3739. Pech, L.L.; Strand, M.R. (2000). Plasmatocyte from the moth Pseudoplusia includens induce apoptosis of granular cells. J. Insect Physiol. 42, 1565–1573.
View at Publisher | View at Google Scholar - Tokura, A.; Fu, G.S.; Sakamoto, M.; Endo, H.; Tanaka, S.; Kikuta, S.; et al. (2014). Factors functioning in nodule melanization of insects and their mechanisms of accumulation in nodules. J. Insect Physiol. 60, 40–49.
View at Publisher | View at Google Scholar
