

Research Article | DOI: https://doi.org/10.31579/2834-5126/007
Dexrazoxane Improves Testicular Tissue Oxidative Stress in Cisplatin-Intoxicated Mice
1School of Medicine,Tehran University of Medical Sciences, Tehran, Iran.
*Corresponding Author: Ojo Olajumoke Omolara, Department of Biochemistry, Ekiti- State University Ado-Ekiti, Nigeria. Email: olajumoke.ojo@eksu.edu.ng
Citation: Ojo O. Omolara, Fajemisin T. Christiana, (2022). Dexrazoxane Improves Testicular Tissue Oxidative Stress in Cisplatin-Intoxicated Mice. J. Clinical trials and clinical research, 1(2); DOI:10.31579/2834-5126/007
Copyright: © 2022, Ojo Olajumoke Omolara. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 21 November 2022 | Accepted: 30 November 2022 | Published: 02 December 2022
Keywords: dexrazoxane; testicular tissue; oxidative stress; cisplatin-intoxicated mice; CPT; dexrazoxane hydrochloride
Abstract
Cisplatin (CPT) compound has shown important biological activities such as antitumor, antituberculous and antimicrobial. However, several researchers have reported the adverse effects of this drug especially on male reproductive system following its administration. Therefore, this study explores the possible ameliorative effect of dexrazoxane against cisplatin-induced testicular tissue using mouse model. Swiss albino mice were randomly selected into three groups (n=5). Group I served as control while group II received 5mg/kg cisplatin and mice in group III were co-treated with 5mg/kg/bwt cisplatin +10mg/kg/bwt dexrazoxane via intraperitoneal injection respectively. Experimental animals from each group were sacrificed at the intervals of 6 hr, 12 hr and 24 hr. The estimation of biochemical and hormonal analyzes by enzyme immunosorbent assay (ELISA) were carried out. Sperm quality and role of apoptosis were also measured in this study. Data were analyzed by Duncan ANOVA at p<0.05. It was observed that CPT leads to significant decrease in level of all the antioxidants measured. CPT leads to significant decrease in the testicular testosterone level and all the antioxidant enzymes measured. Cisplatin leads to depletion in the number of spermatozoa cells and increased aberrant spermatozoa cells structure. However, recovery was seen in the group co-treated with dexrazoxane. This study gives insight on the adverse effects of cisplatin compound on male reproductive system also shows that melatonin remarkably improved the deleterious effect seen in the all parameters tested.
Introduction
Cisplatin is a platinum based chemotherapy agent. It has been used for treatment of numerous human cancers including testicular cancers. Cisplatin mode of action has been linked to its ability to crosslink with the purine bases on the DNA; interfering with the DNA repair mechanisms, causing DNA damage, and subsequently inducing apoptosis in cancer cells [1-3]. However, due to numerous undesirable side effects especially in younger patients and fetus, its use was restricted but combination therapies of cisplatin with other drugs have been highly encouraged to overcome drug-resistance and reduce toxicity. Dexrazoxane hydrochloride is a cardioprotective agent used to protect the heart against the cardiotoxic side effects of chemotherapeutic drugs such as anthracyclines [4], daunorubicin or doxorubicin or other chemotherapeutic agents [5]. Based on the available data legally binding the decision to implement dexrazoxane in order to allow children to receive dexrazoxane as primary cardioprotection against anthracyclineinduced cardiotoxicity without reducing anthracycline activity and without enhancing secondary malignancies [6], It was therefore speculated that dexrazoxane could be used for further investigation to synthesize new drugs [7]. Hence, this study, explore dexrazoxane use in the ameliorating the deleterious effect of cisplatin in testicular tissue dysfunction.
2.0. Methods
2.1 Ethnical Approval
The studies were conducted in accordance with the standards and permission established by The Ethics Committee of Animal, Ekiti State University Ado-Ekiti, Nigeria. Male Swiss-albino mice were housed in room at 22 ± 2°C with 40% relative humidity and with a 12-hr light ± dark cycle. They were fed with a standard rat chow and tap water ad libitum.
2.2 Experimental Procedure
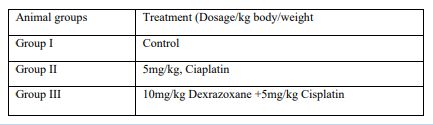
Male Swiss-albino mice used for this study were weighed. The animals were randomly divided into three group I-III and treated as shown in the table below.
Dosing
2.3 Testicular Testosterone (T) and Luteinizing hormone (LH) concentrations.
The testicular testosterone and luteinizing hormone levels in three mice from each group were measured. Briefly, testicular proteins were extracted with phosphate buffer (50 mM, pH 7.4) and centrifuged at 10,000 g for 20 min. The supernatant was used to estimate T and LH levels using ELISA, and were expressed in ng/ml.
2.5 Biochemical Estimations study
Testicular tissues from each mouse were stored at -20oC for different biochemical assays Protein quantity was estimated according to Lowry’s method. 10% tissue homogenates (w/v) were prepared in chilled 100 mMTris-HCL buffer (pH 7.4). The values were expressed per mg of protein.
2.6 GSH Determination
Tetsis tissues were homogenized in 10 ml TCA (trichloroacetic acid) which is at the rate of 10%, and then centrifuged at +4 °C for 15 minutes. Afterwards, 0,5 ml of supernatant was taken, and mixed with 0,3 M 2 ml Na2HPO4. The mixture was thoroughly vortexed. This mixture was vortexed by the addition of 0,2 ml DTBN (Dithiobisnitrobenzene: prepared by dissolving in 1% sodium citrate). Absorbance was measured at 412 nm.
2.7. Measurement of reactive oxygen species (ROS) level
The ROS assay was performed by the method of Hayashi., et al. (2017). In brief, 50 µl of testicular tissue homogenate and 1400 µl sodium acetate buffer was transferred to a cuvette. After then, 1000 ul of reagent mixture (N, N-diethyl paraphenylenediamine 6 mg/ml with 4.37 µM of ferrous sulfate dissolved in 0.1M sodium acetate buffer pH- 4.8) was added at 37°C for 5 minutes. The absorbance was measured at 505 nm using spectrophotometer [8].
2.8. Measurement of MDA
Testes tissues were homogenized in 10 ml TCA (trichloroacetic acid) which is at the rate of 10%, and then centrifuged at +4°C for 15 minutes. 750μl of the supernatant which was obtained was mixed with 0.67% TBA (thiobarbituric acid) in a ratio of 1:1. Afterwards, the solution was left in the water bath for 15 minutes. Finally, the absorbance was measured spectrophotometrically at 535 nm.
2.9. Superoxide dismutase (SOD) activity
Superoxide dismutase (SOD) activity was assayed by a spectrophotometric method. Assay mixture containing sodium pyrophosphate buffer (pH 8.3, 0.052M), phenazine methosulfate (186 µM), nitroblue tetrazolium (300 µM) and NADH (780 µM) were diluted with appropriate enzyme in total volume of 3 ml. The mixture was incubated at 37oC for 90 sec and reaction was stopped by addition of glacial acetic acid. The reaction mixture was mixed vigorously by adding n-butanol and allowed to stand for 10 min before the collection of butanol layer. The intensity of chromogen in butanol was measured at 520 nm.
2.10. Sperm Parameters
Caudal epididymidis was removed from each mouse and cleaned off from the epididymal fat pad, and minced in a pre-warmed Petri dish containing 500 µl phosphate buffer saline solutions (PBS, pH 7.4) at 37oC. Sperm motility was estimated and expressed as percentage incidence [4]. For sperm count, an aliquot of this suspension was charged into the Neubauer’s counting chamber and the spermatozoa were counted under light microscope. Total sperm count was calculated as the average of the spermatozoa count (N) in each chamber X multiplication factor (106) X dilution factor and was expressed in millions/ml. The sperm morphology was also evaluated [9].
2.11. Capases Estimation
Briefly, 1ml of assay buffer (20mM HEPES, 10% glycerol, 1M DTT, and 14ml of n-acetyl-DEVD-AMC/ml of buffer), and 50ml of sample were added to a microcentrifuge tube and protected from the light. Samples were incubated at 37°C for 60 mins after which fluorescence was measured on a spectrofluorometer with an excitation wavelength of 380nm and an emission wavelength of 440nm.
2.12. Statistical Analysis
All data were expressed as mean ±standard error of the mean (SEM) and analyzed by one-way ANOVA followed by Duncan’s multiple comparison test using SPSS software version 22 (SPSS Inc., Chicago, Illinois). p<0> were considered statically significant.
3.1 The effect on Reactive oxygen species
A significant increase was observed in CPT-treated group compared to the control group (P<0>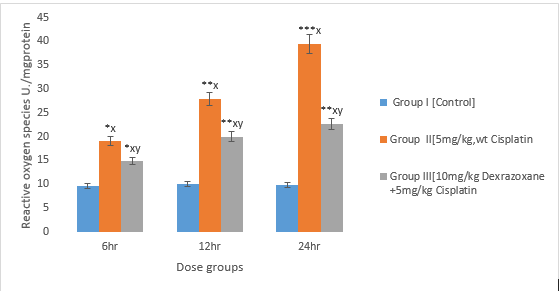
3.2 The effect on Malonialdehyde
There was an increase observed in CPT-treated group compared to the control group (P<0>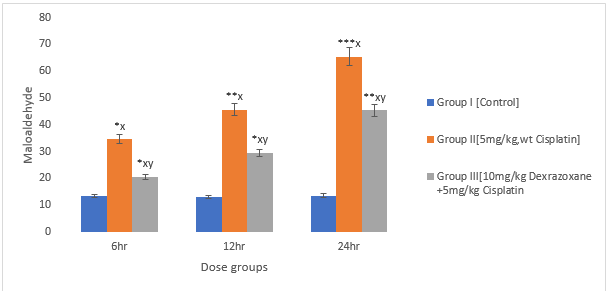
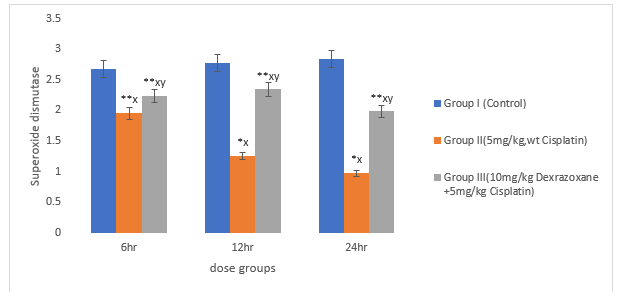
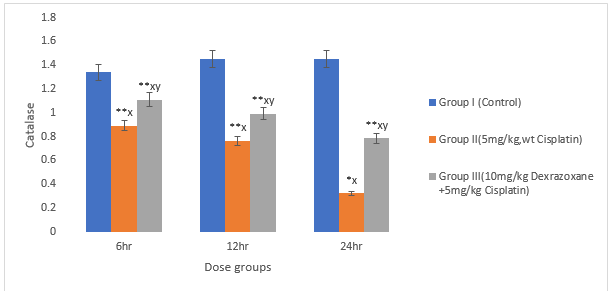
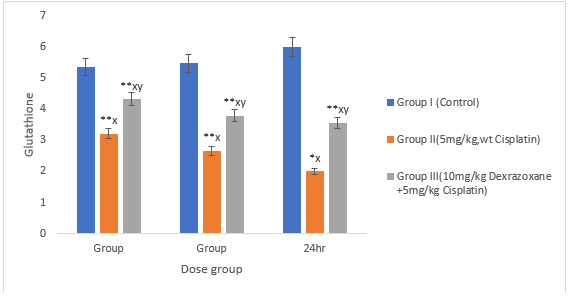
3.3 Effect on antioxidant status
There was significant decrease in the superoxide dismustase, catalase and glutathione when the mice were treated with CPT (P>0.01) compared to the control. When co-treated with DZX+CPT there was significant recovery compared to the CPT-treated group (figures 3, 4 & 5).
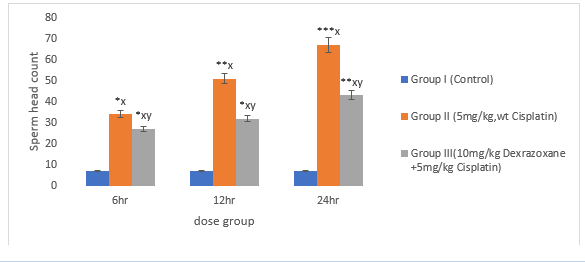
3.6 Effect of on Sperm Head Count
There was significant increase in CPT-treated group compared to the control (P<0>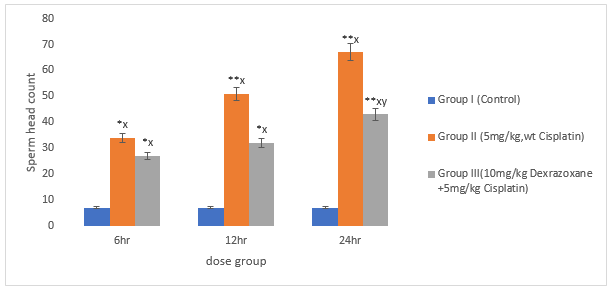
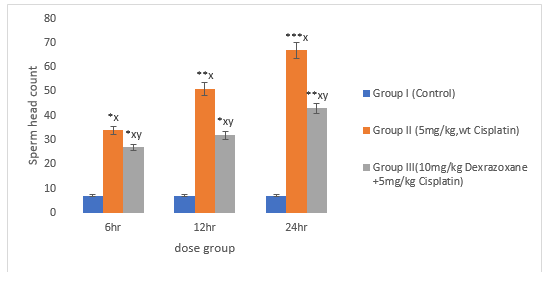
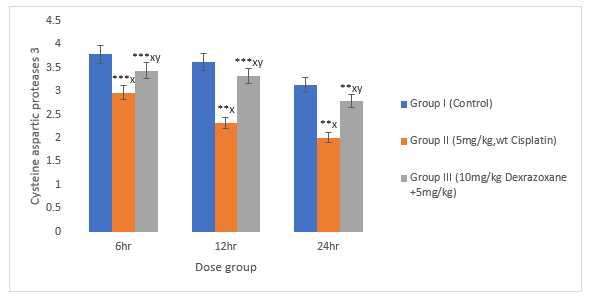
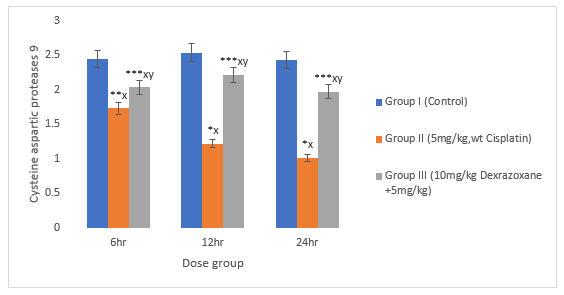
3.9 Effect on Caspase 3 and 9
There was significant decrease in CPT-treated group compared to the control group (P>0.01). When treated with DZX+CPT group, there is significant increase in cysteine aspartic proteases compared to the CPT-treated group (figures 9 & 10).
Discussion
Cisplatin induces excessive ROS production that disrupts physiological balance of redox and anti-oxidation activities [10] It had been previously shown that both in-vitro and in-vivo cisplatin compromised mitochondria total antioxidant capacity, and as a consequence, it led to imbalanced mitochondria redox processes and caspase 3-associated apoptosis [11,12]. Other studies also showed cisplatin induced dose-dependent testicular damage, ROS generation and ER stress in rat testis. It was observed from the experiment that cisplatin significantly increases ROS production. Cancer cells exhibit greater reactive oxygen species stress that normal cells do, partly due to oncogenic stimulation, increased metabolic activity and mitochondrial malfunction. Oxidative stress is the one of the most important mechanisms involved in cisplatin toxicity. The mitochondrion is the primary target for cisplatin induced oxidative stress, resulting in loss of mitochondrial protein sulfhydrl group, calcium uptake inhibition and reduction of mitochondrial membrane potential [13]. Exposure to oxidative stress can upset regular biological functions. Cisplatin can also induce reactive oxygen species that trigger cells death besides DNA damage.
Cells death occurs upon immediate activation of numerous signaling pathways, whereas the definite pathways depend on the cells. The formation of reactive oxygen species depends on the concentration of cis-diamminedichloro platinum (II) and the length of exposure [14]. The intracellular redox homeostasis is maintained by the thiol group (-SH) containing molecules. Under certain conditions a thiol group may lead to the formation of thiyl radicals that in turn can interact with oxygen, therefore generating reactive oxygen species [15].
A decrease was observed in catalase activity but an increased level of malonialdehyde. Relieving effect was observed when dexrazoxane was added. Appraisal of the results together with the literature suggests that cisplatin treatment disrupts both enzymatic antioxidant activities. Mitochondrial enzymes, glutathione and superoxide dismutase have been correlated with cellular anti-oxidation activity [16,17].
Cisplatin has been reported to exert its injurious effects on testis by increasing MDA levels and eliminating antioxidants [18,19]. Also, cisplatin-related injury including, depletion of germ cells, atrophy of seminiferous tubules and dropout of sertoli cells within lumen in testis had been been reported, this study shows possible potential of dexrazoxane protective effect against cisplatin-induced injury. It is possible that dexrazoxane protects testicles by many other mechanisms, because it has been recently pointed out that this bioflavonoid prevents chronic cadmium-induced nephrotoxicity by overexpressing endothelial nitric oxide synthase and cyclooxygenase-2 in rats [20].
Sperm motility and morphology were also affected by cisplatin, and this negative effect has been reversed by dexrazoxane. It is possible that the increased epididymal sperm quality might be a result of the antioxidant activity of dexrazoxane on the epididymis [21]. The epididymis, serving important functions in the transportation, maturation and storage of sperm cells, during which period the spermatozoa develop motility, it has been shown to be enlarged in dexrazoxane-treated mice [22]. The epididymis also protects spermatozoa from oxidative injury by encouraging scavengers of reactive oxygen species [23]. Testicular oxidative stress is known to play a role in a number of conditions detrimental to male fertility. It is generally accepted that the increased lipid peroxidation is one of the toxic manifestations of cisplatin administration in testis. The reduction in sperm motility and sperm morphology in cisplatin-treated rats may be caused by lipid peroxidation of unsaturated fatty acids in the sperm plasma membrane, causing in a loss of its fluidity and function. Previous studies shown that, the treatment of male rats with cisplatin resulted in a significant increase in testicular tissue levels of MDA [24]. In the present study, sperm quality, as evaluated by the epididymal sperm motility and sperm viability, was found to be improved by the dexraxozane treatment. Conclusively, cisplatin is considered as cytotoxic drug which kills cancer cells by damaging DNA, inhibiting DNA synthesis and mitosis, and inducing apoptotic cell death. Several molecular mechanisms of action including induction of oxidative stress as characterized by reactive oxygen species production and lipid peroxidation as being linked to the administration of this anticancer agent however its combination with dexrazoxane which target multiple mechanisms, such as reducing cisplatin uptake and inflammation, may offer the best chance for clinically meaningful prevention of cisplatin toxicity if further explored.
References
- Fischer J, Ganellin CR (2006). Analogue-based Drug Discovery. John Wiley & Sons. p. 513. ISBN 9783527607495. Archived from the original on 20 December 2016.
View at Publisher | View at Google Scholar - Lipshultz SE, Rifai N, Dalton VM, Levy DE, Silverman LB, Lipsitz SR, et al. (2004). The effect of dexrazoxane on myocardial injury in doxorubicin-treated children with acute lymphoblastic leukemia. The New England Journal of Medicine. 351 (2): 145–153.
View at Publisher | View at Google Scholar - Bjelogrlic SK, Radic J, Radulovic S, Jokanovic M, Jovic V (2007). Effects of dexrazoxane and amifostine on evolution of Doxorubicin cardiomyopathy in vivo. Experimental Biology and Medicine. 232 (11): 1414–1424.
View at Publisher | View at Google Scholar - Salzer WL, Devidas M, Carroll WL, Winick N, Pullen J, Hunger SP, Camitta BA (2010). Long-term results of the pediatric oncology group studies for childhood acute lymphoblastic leukemia 1984-2001: a report from the children's oncology group. Leukemia. 24 (2): 355–370.
View at Publisher | View at Google Scholar - Kane RC, McGuinn WD, Dagher R, Justice R, Pazdur R (2008). Dexrazoxane (Totect): FDA review and approval for the treatment of accidental extravasation following intravenous anthracycline chemotherapy. The Oncologist. 13 (4): 445–450.
View at Publisher | View at Google Scholar - Jones RL (2008). Utility of dexrazoxane for the reduction of anthracycline-induced cardiotoxicity. Expert Review of Cardiovascular Therapy. 6 (10): 1311–1317.
View at Publisher | View at Google Scholar - Loyevsky M, Sacci JB, Boehme P, Weglicki W, John C, Gordeuk VR (1999). Plasmodium falciparum and Plasmodium yoelii: effect of the iron chelation prodrug dexrazoxane on in vitro cultures. Experimental Parasitology. 91 (2): 105–114.
View at Publisher | View at Google Scholar - Olajumoke Omolara Ojo and Yemisi Rufina Alli Smith. (2020). Adverse Effect of Dactinomycin on Reactive Oxygen Species and Important Steriodogenic Enzymes in Male Mice. EC Pharmacology and Toxicology 8(3):1-9.
View at Publisher | View at Google Scholar - Ojo OO, Bhadauria S, Rath SK. (2013). Dose Dependent Adverse Effects of Salinomycin on Male Reproductive Organs and Fertility in Mice. PLOS One. 2013; 2(6):133-140, e69086.
View at Publisher | View at Google Scholar - Liu, H.T.; Wang, T.E.; Hsu, Y.T.; Chou, C.C.; Huang, K.H.; Hsu, C.C.; Liang, H.J.; Chang, H.W.; Lee. T.H.; Tsai, P.S. Nanoparticulated Honokiol Mitigates Cisplatin-induced Chronic Kidney Injury by Maintaining Mitochondrial Antioxidation Capacity and Reducing Caspase-3-Associated Cellular Apoptosis. Antioxidant 2019, 8, 466.
View at Publisher | View at Google Scholar - Wang, T.J.; Liu, H.Y.; Lai, Y.H.; Jan, T.R.; Nomura, N.; Chang, H.W.; Chou, C.C.; Lee, Y.J.; Tsai, P.J. (2018). Honokiol, a polyphenol Natural Compound Attenuates Cisplatin-induced Acute Cytotoxicity in Renal Epithelial Cells Through Cellular Oxidative Stress and Cytoskeleton Modulations. Front. Pharmacol. 2018, 9, 357.
View at Publisher | View at Google Scholar - Brozovic A, Ambriovic-Ristov A, Osmak M. (2010). The relationship between Cisplatin-induced reactive oxygen species, glutathione and BCl-2 and resistance to cisplatin. Crit Rev. Toxicol. 2010; 40:347-359.
View at Publisher | View at Google Scholar - Desoize B. (2002). Cancer and metals and metals compounds; part 1-carcinogenesis. Crit Rev. oncol. Hematol. 2002; 42:1-3.
View at Publisher | View at Google Scholar - Miyamoto, Y.; Koh, Y.H.; Park, Y.S.; Fujiwara, N.; Sakiyama, H.; Misonou, Y.; Ookawara, T.; Suzuki, K.; Honke, K.; Taniguchi, N. (2003). Oxidative stress caused by inactivation of glutathione peroxidase and adaptive responses. Biol. Chem. 2003, 384, 567-574.
View at Publisher | View at Google Scholar - Martindale JL, Holbrook NJ. (2002). Cellular response to oxidative stress: signaling for suicide and survival. J. Cell Physiol. 2002; 192:1-15.
View at Publisher | View at Google Scholar - Hampton MB, Orrenius S. (1997). Dual regulation of caspase activity by hydrogen peroxide: implication of apoptosis. FEBS Lett.1997; 414:552-556.
View at Publisher | View at Google Scholar - Silici S, Ekmekcioglu O, Kanbur M, Deniz K (2011). The protective effect of royal jelly against cisplatin-induced renal oxidative stress in rats. World j Urol 29:127-132.
View at Publisher | View at Google Scholar - Salem EA, Salem NA, Maarouf AM, Serefoglu EC, Hellstorm WJG (2012). Selenium and lycopene attenuate cisplatin-induced testicular toxicity associated with oxidative stress in wistar rats. Urology 79: 1184.E 1-6.
View at Publisher | View at Google Scholar - Kim JC, Kim KH, Chung MK (1999). Testicular cytotoxicity of DA-125, a new anthracycline anti-cancer agent, in rats. Reprod Toxicol 13: 391-397.
View at Publisher | View at Google Scholar - Atessahin A, Sahna E, Turk G, Ceribasi AO, Yilmaz S, Yuce A, Bulmus O (2006). Chemoprotective effect of melatonin against cisplatin-induced testicular toxicity in rats. J Pineal Res 41:21-27.
View at Publisher | View at Google Scholar - Sukhotnik I., et al. (2013). Methotrexate induces germ cell apoptosis and impair spermatogenesis in a rat. Pediatric Surgery International 29.2 (2013): 179-184.
View at Publisher | View at Google Scholar - Morales Al, vincente-sanchezc, jerkic M, Santiago JM, Sanchez- Gonzalez PD, perez-Barricanal F, Lopez-Noova JM (2006). Effect of quercetin on metallothionein, nitric oxide synthases and cyclooxygenase-2 expression on experimental chronic cadmium nephrotoxicity in rats. Toxicol Appl Pharmacol 210: 128-135.
View at Publisher | View at Google Scholar - Zini A, Schlegel PN (1997). Identification and characterization of antioxidant enzymes mRNAs in the rat epididymis. Int Androl 20:86-91.
View at Publisher | View at Google Scholar - Taepongsorat L, Tangpraprutgul P, Kitana N, Malaivijitnond S. (2008). Stimulating effects of quercetin on sperm quality and reproductive organs in adult male rats. Asian j Androl 10:249-258.
View at Publisher | View at Google Scholar
