

Review Article | DOI: https://doi.org/10.31579/2834-8761/105
Advances in Cervical Cancer Therapeutics: HPV Vaccine Design, Immunological Mechanisms, and Future Perspectives
Department of Zoology, Deen Dayal Upadhyay Gorakhpur University, Gorakhpur, India.
*Corresponding Author: Ravi Kant Upadhyay, Department of Zoology, Deen Dayal Upadhyay Gorakhpur University, Gorakhpur, India.
Citation: Arfiya Fatima, Ravi K. Upadhyay, (2026), Advances in Cervical Cancer Therapeutics: HPV Vaccine Design, Immunological Mechanisms, and Future Perspectives, Clinical Endocrinology and Metabolism, 5(2); Doi:10.31579/2834-8761/105
Copyright: © 2026, Ravi Kant Upadhyay. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 17 February 2026 | Accepted: 04 March 2026 | Published: 11 March 2026
Keywords: cervical cancer; human papillomavirus; hpv vaccine; prophylactic vaccine; therapeutic vaccine; immunotherapy; immune checkpoint inhibitors; vaccine design
Abstract
This article presents various causes of cervical cancer, immunological methods and vaccines use for cervical cancer. It also sketches out various risk factors and diagnostic methods. HPV biology and immunopathogenesis, conventional and emerging treatment strategies, and the current landscape of prophylactic and therapeutic HPV vaccines. Particular attention is given to DNA-, RNA-, peptide-, viral vector-, and cell-based vaccine platforms, as well as nanoparticle-assisted delivery systems and combination strategies with immune checkpoint inhibitors. We also highlight implementation challenges, the World Health Organization cervical cancer elimination strategy, and future opportunities in personalized immunotherapy, biomarker-guided management, and equitable vaccine deployment. A multidisciplinary approach integrating vaccination, screening, early diagnosis, and innovative therapeutics will be essential for reducing disease burden and advancing cervical cancer control. Among therapeutic HPV vaccines, all these induce strong cell-mediated immunity, especially cytotoxic CD8+ T-cell responses, against the viral antigens constitutively expressed in infected and transformed cells. Combination therapy may improve antigen release, dendritic cell activation, intra tumoral T-cell recruitment, and reversal of immunosuppression.
Introduction
Cervical cancer is the fourth most common cancer among women globally and remains one of the most unequally distributed malignancies, with the highest burden falling on low- and middle-income countries [1,2]. According to the World Health Organization (WHO), nearly 94% of cervical cancer deaths in 2022 occurred in low- and middle-income settings, where access to vaccination, screening, and timely treatment is often limited [3]. The WHO global elimination framework has therefore prioritized cervical cancer as a preventable cancer for coordinated public health action through its 90–70–90 targets: 90% HPV vaccination coverage in girls by age 15 years, 70% screening coverage by ages 35 and 45 years, and 90% treatment access for women with precancer or invasive disease [3]. Persistent infection with high-risk human papillomavirus (hrHPV), particularly HPV-16 and HPV-18, is the dominant etiological factor in the development of cervical cancer [4,5]. These viral types encode the oncoproteins E6 and E7, which disrupt critical tumor suppressor pathways involving p53 and pRb, thereby enabling cellular immortalization, genomic instability, and malignant progression [5,6]. While most HPV infections are transient and cleared by host immunity, persistent infection can lead to cervical intraepithelial neoplasia (CIN) and eventually invasive carcinoma [4,7]. Doorbar et al. and Graham have established the central role of persistent hrHPV infection in progression from precancerous lesions to invasive disease [4,7]. Over the last two decades, prophylactic HPV vaccines have transformed prevention strategies and have demonstrated strong efficacy against vaccine-type HPV infection and precancerous lesions [8,9]. However, these vaccines do not eliminate pre-existing infection or established neoplastic lesions [8]. This limitation has driven increasing interest in therapeutic vaccines designed to induce cell-mediated immune responses against infected or transformed cells, primarily through targeting the HPV E6 and E7 antigens [10,11]. Recent advances in immunotherapy, viral vectors, nucleic acid vaccines, nanotechnology, and tumor immunology have expanded the therapeutic landscape of cervical cancer beyond surgery, radiotherapy, and chemotherapy [11–13]. This review discusses current advances in cervical cancer therapeutics with a particular focus on HPV vaccine design, immunological mechanisms, and future directions. It also highlights major implementation barriers, translational opportunities, and the need for integrated prevention and treatment strategies.
Methods
This narrative review was prepared through literature searches of PubMed/MEDLINE, Google Scholar, and major public health and regulatory resources, including WHO and the United States Food and Drug Administration (FDA). Search terms included “cervical cancer”, “human papillomavirus”, “HPV vaccine”, “therapeutic vaccine”, “prophylactic vaccine”, “E6”, “E7”, “immune checkpoint inhibitors”, “DNA vaccine”, “mRNA vaccine”, “nanoparticles”, and “HPV-negative cervical cancer”. Priority was given to recent peer-reviewed reviews, original studies, clinical trial discussions, and authoritative global guidance documents published within the last 10 years, while seminal earlier references were included where necessary for biological context. Information was synthesized narratively according to thematic relevance. Because this is a narrative review, a formal risk-of-bias assessment was not performed.
3. Global burden, Epidemiology, and risk Factors of Cervical Cancer
Cervical cancer remains a significant global public health challenge and ranks as the fourth most common cancer among women worldwide. According to estimates from GLOBOCAN 2022, approximately 600,000 new cases and over 340,000 deaths occur annually, with nearly 85–90% of these deaths reported in low- and middle-income countries (LMICs) [11,12]. This disproportionate burden is primarily driven by limited access to effective screening programs, inadequate healthcare infrastructure, and low uptake of human papillomavirus (HPV) vaccination [12,13]. The epidemiology of cervical cancer shows marked geographic variation. The highest incidence and mortality rates are observed in sub-Saharan Africa, South Asia, and parts of Latin America, whereas developed regions have experienced substantial declines due to organized screening programs, early detection, and widespread vaccination [11,14]. These disparities highlight the critical role of healthcare accessibility and preventive strategies in reducing disease burden.
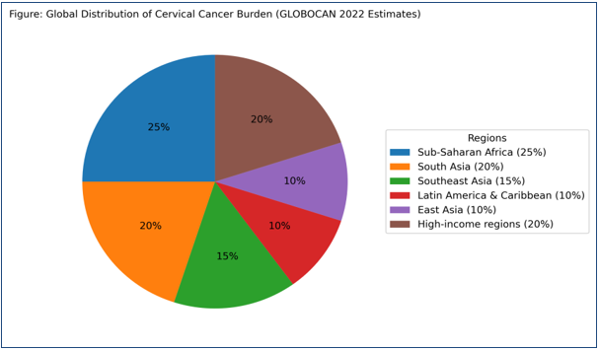
Figure 1: shows the global distribution of the cancer burden (GLOBOCAN estimates).
Persistent infection with high-risk HPV (hrHPV), particularly genotypes HPV-16 and HPV-18, is the primary etiological factor in cervical carcinogenesis [15,16]. However, progression from infection to invasive cancer is multifactorial. Several behavioral, biological, and socio-economic cofactors contribute to disease development, including early age at first sexual intercourse, multiple sexual partners, long-term use of oral contraceptives, smoking, high parity, and immunosuppression, especially in individuals with human immunodeficiency virus (HIV) infection [13,17]. Smoking plays a particularly important role by introducing carcinogenic compounds that impair local immune responses and induce genetic damage in cervical epithelial cells [18]. Additionally, emerging evidence suggests that alterations in cervicovaginal microbiota, particularly the depletion of Lactobacillus-dominant communities, may promote chronic inflammation and facilitate HPV persistence, although further longitudinal studies are required to establish causality [19]. Socio-economic determinants such as poverty, lack of awareness, cultural barriers, and poor access to healthcare services further exacerbate the burden of cervical cancer in resource-limited settings [12,13]. Despite advances in prevention through HPV vaccination and screening, global projections indicate that the incidence and mortality may continue to rise unless equitable and large-scale interventions are implemented [11]. Although the majority of cervical cancers are HPV-associated, a small subset is HPV-negative. These tumors often exhibit distinct molecular and pathological characteristics and may pose diagnostic challenges, as they are not detectable through HPV-based screening approaches. Their pathogenesis remains incompletely understood and warrants further investigation [20]. Overall, cervical cancer represents a largely preventable malignancy; however, persistent global inequalities in prevention, early detection, and treatment remain the primary drivers of high mortality [12].
4. Immunopathogenesis of Cervical cancer
4.1 HPV Structure and Life Cycle
Human papillomaviruses (HPVs) are small (50–60 nm), non-enveloped, double-stranded DNA viruses belonging to the Papillomaviridae family [21,22]. The viral capsid is composed of 72 capsomeres formed by pentameric L1 proteins along with the minor capsid protein L2. The circular genome (~7–8 kb) is functionally divided into early (E1–E7) genes, late (L1 and L2) structural genes, and a non-coding long control region (LCR), which regulates viral replication and transcription [21].
HPVs exhibit strict species specificity and pronounced epithelial tropism, infecting basal keratinocytes of stratified squamous epithelium. Infection is initiated through micro-abrasions in the cervical epithelium, allowing viral entry into the basal layer [23]. In the early phase, viral genomes are maintained episomally at low copy numbers, mediated by the coordinated action of E1 (helicase) and E2 (transcriptional regulator) [24]. As infected basal cells proliferate and differentiate, they migrate toward the suprabasal layers, where the virus exploits host differentiation machinery to amplify its genome. This differentiation-dependent life cycle results in high-level viral replication, expression of late genes (L1, L2), and assembly of progeny virions, which are released without causing cell lysis [25]. This non-cytolytic nature of infection contributes to minimal immune activation and facilitates viral persistence [22]. Among more than 200 identified HPV genotypes, high-risk types—particularly HPV-16 and HPV-18—are responsible for approximately 70% of cervical cancer cases [16,26]. While most HPV infections are transient and cleared by the immune system, persistent infection with high-risk types is a prerequisite for malignant transformation [23].
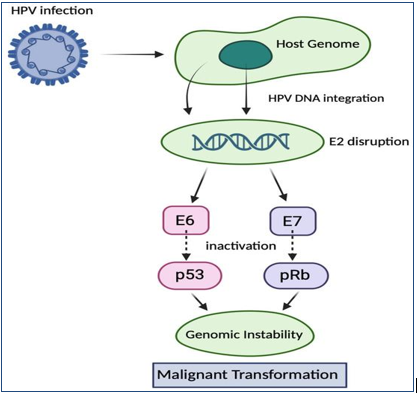
Figure 2: shows HPV infection and malignant transformation inside the host.
4.2 HPV Integration and Immune Evasion
• HPV Pathogenesis and Disease Progression
HPV infection primarily targets mucosal epithelial cells, and in most individuals, it is effectively cleared by host immune responses. However, in a subset of cases, persistent infection leads to the development of precancerous lesions known as cervical intraepithelial neoplasia (CIN). These lesions are graded as CIN1 (low-grade), CIN2, and CIN3 (high-grade) [27]. While CIN1 lesions frequently regress due to immune-mediated clearance, persistent infection can drive progression to high-grade lesions and eventually invasive cervical carcinoma [23,27].
• HPV Integration and Oncogenic Transformation
A critical event in cervical carcinogenesis is the integration of HPV DNA into the host genome. This process often disrupts the viral E2 gene, a negative regulator of E6 and E7 expression, resulting in their constitutive overexpression [28].
The viral oncoproteins E6 and E7 play central roles in tumorigenesis:
• E6 protein promotes ubiquitin-mediated degradation of p53 via E6-associated protein (E6-AP), thereby inhibiting apoptosis, impairing DNA repair mechanisms, and inducing telomerase activation, leading to cellular immortalization [29].
• E7 protein binds to and inactivates the retinoblastoma (pRb) protein, releasing E2F transcription factors and driving uncontrolled cell cycle progression. E7 also inhibits cyclin-dependent kinase inhibitors such as p21 and p27 [30].
These interactions result in genomic instability, accumulation of mutations, and deregulated cellular proliferation, ultimately promoting malignant transformation [28–30].
• Immune Evasion Mechanisms
HPV has evolved sophisticated mechanisms to evade host immune surveillance, including:
• Downregulation of MHC class I molecules, impairing antigen presentation
• Suppression of interferon signaling pathways
• Lack of cytolysis, leading to minimal inflammation
• Modulation of the local immune microenvironment
These immune evasion strategies allow persistent infection and tumor progression [31,32]. Cofactors such as HIV infection, smoking, and chronic inflammation further weaken immune responses and enhance viral persistence [13,18]. Smoking, in particular, contributes to DNA damage and accumulation of carcinogens in cervical mucus, thereby facilitating viral integration and oncogenesis [18].
• Molecular Pathways and Cellular Reprogramming
HPV oncoproteins (E5, E6, E7) dysregulate multiple signaling pathways, including p53, Rb, Wnt/β-catenin, MAPK, PI3K/Akt/mTOR, and STAT pathways [33]. These alterations collectively reprogram host cellular machinery, promoting proliferation, inhibiting apoptosis, and enabling immune escape.
Epigenetic modifications, such as aberrant DNA methylation, along with biomarkers like p16 overexpression, are widely used to detect HPV-associated transformation and disease progression [34].
• Tumor Suppressor Restoration and Therapeutic Insights
Given the central role of p53 inactivation, restoration strategies such as p53 gene therapy (e.g., recombinant adenoviral p53 delivery) have been explored. Clinical studies have demonstrated improved local tumor control when combined with radiotherapy, although further optimization and large-scale validation are required [35].
5. HPV vaccines in prevention and treatment
Human papillomavirus (HPV) is one of the main causes of cervical cancer and other HPV-related diseases [36,37]. Vaccination has become an important way to reduce the spread of HPV infection and lower the risk of disease development [38]. At present, three licensed preventive HPV vaccines are available: bivalent, quadrivalent, and nonavalent vaccines [39]. These vaccines are made from L1 virus-like particles and have greatly helped in reducing HPV infections worldwide. It is estimated that vaccination can prevent nearly 90% of HPV infections and related diseases [40]. However, these vaccines do not treat infections or lesions that are already present [41]. For this reason, therapeutic HPV vaccines are being developed. These vaccines mainly target the E6 and E7 oncoproteins of HPV and work by activating cellular immunity instead of producing neutralizing antibodies [42]. Therefore, therapeutic vaccines are considered a promising approach for clearing existing HPV infection and HPV-related lesions [43].
5.1 Prophylactic vaccines
Prophylactic human papillomavirus (HPV) vaccination is one of the most effective strategies for preventing cervical cancer (CC) [44,45]. These vaccines are recommended for both females and males, ideally between the ages of 9–14 years, typically administered in a two-dose schedule [44,45]. They primarily target high-risk HPV types responsible for the majority of cervical cancer cases [46]. Currently available HPV vaccines include bivalent, quadrivalent, and nonavalent formulations. The bivalent vaccine (Cervarix) targets HPV types 16 and 18, which are responsible for most cervical cancer cases [46]. The quadrivalent vaccine (Gardasil) protects against HPV types 6, 11, 16, and 18, while the nonavalent vaccine (Gardasil 9) provides broader protection against nine HPV types, including 6, 11, 16, 18, 31, 33, 45, 52, and 58 [47–49].
These vaccines are based on virus-like particles (VLPs) composed of the L1 capsid protein. They are highly immunogenic and induce neutralizing antibodies that prevent initial HPV infection. Since they do not contain viral DNA, they are non-infectious [48,50]. According to World Health Organization (WHO) reports (2024–2025), several HPV vaccines have been prequalified, including Cervarix, Gardasil, Gardasil 9, Cecolin, and Walrinvax, while Cervavac has been licensed for use in India [45,51]. WHO has also endorsed single-dose vaccination strategies for certain HPV vaccines under evidence-based conditions to improve accessibility and coverage [51]. Despite their strong preventive efficacy, HPV vaccines do not have therapeutic effects. They cannot eliminate existing HPV infections, persistent cervical intraepithelial neoplasia (CIN), or invasive cervical cancer [41,50]. This is because viral capsid proteins (L1 and L2) are not sufficiently expressed in transformed basal cells, making them ineffective targets for immune clearance in established infections [42]. Therefore, regular cervical cancer screening remains essential, even among vaccinated individuals, to ensure early detection and proper management [44].
5.2 Preventive HPV Vaccines and the Need for Therapeutic Vaccines
The identification of human papillomavirus (HPV) as a major etiological agent of cervical cancer and other HPV-associated malignancies has led to the development of effective immunization strategies aimed at preventing infection and disease progression [36,37]. Prophylactic HPV vaccines are designed to induce protective humoral immunity by targeting the viral capsid proteins L1 and, in some cases, L2. These vaccines utilize virus-like particles (VLPs), which are non-infectious structures lacking viral DNA but closely resemble native virions in terms of structure and immunogenicity, thereby eliciting strong neutralizing antibody responses [48–50]. Currently available prophylactic vaccines are based on recombinant L1 VLPs and have demonstrated high efficacy in preventing infections caused by high-risk HPV types, particularly those associated with cervical cancer [47,49]. These vaccines generate type-specific neutralizing IgG antibodies, and their protective efficacy is highly dependent on the conformational integrity of the L1 protein [50]. Preclinical studies have shown that only structurally intact VLPs can induce protective immunity, highlighting the importance of conformational epitopes in vaccine design [50]. Despite their success, these vaccines do not eliminate pre-existing HPV infections or established lesions, as they primarily act by preventing viral entry rather than clearing infected cells [41,47]. One key limitation of prophylactic vaccines is that the late capsid proteins (L1 and L2) are not expressed in basal epithelial cells harboring persistent infection or in transformed cancer cells. As a result, these proteins are not suitable targets for therapeutic intervention [42]. In contrast, the viral oncoproteins E6 and E7 are consistently expressed in HPV-infected and transformed cells and are essential for maintaining the malignant phenotype, making them ideal targets for therapeutic vaccine development [42,43]. Therapeutic HPV vaccines aim to overcome these limitations by inducing robust cell-mediated immune responses, particularly cytotoxic T lymphocyte (CTL) activity, against infected or transformed cells [43]. Among the various platforms under development, DNA vaccines have gained significant attention due to their safety, stability, ease of large-scale production, and ability to induce sustained immune responses [52]. Unlike RNA vaccines, DNA vaccines are more stable, cost-effective for storage and transport, and allow repeated administration without significant toxicity. Additionally, multiple plasmids encoding different antigens can be combined into a single formulation, enhancing their immunogenic potential [52]. Despite the availability of effective prophylactic vaccines, HPV infection and cervical cancer remain prevalent globally, largely due to unequal vaccine coverage and limited access in low- and middle-income countries [51]. Therefore, there is a critical need for therapeutic strategies capable of treating established infections and associated lesions. Therapeutic HPV vaccines, by targeting continuously expressed viral antigens such as E6 and E7, offer a promising approach for eliminating infected cells and reducing disease burden. Consequently, the integration of both preventive and therapeutic vaccination strategies is essential for comprehensive control of cervical cancer [43,51].
| Brand name | Valency | Target-HPV Types |
| Cervarix | bivalent | 16, 18 |
| Cecolin | bivalent | 16, 18 |
| Walrinvax | bivalent | 16, 18 |
| Gardasil | quadrivalent | 16, 18 |
| Cervavax | quadrivalent | 16, 18, 6, 11 |
| Gardasil9 | nonvalent | 16, 18, 31, 33, 45, 52, 58, 6, 11 |
Table 1: WHO-licensed HPV vaccines 2024
5.3 Therapeutic vaccines
In contrast to prophylactic vaccines, which primarily induce neutralizing antibody responses, therapeutic HPV vaccines are designed to generate strong cell-mediated immunity, particularly cytotoxic CD8⁺ T-cell responses against infected and transformed cells [53,54]. The major targets of these vaccines are the viral oncoproteins E6 and E7, as they play a central role in HPV-induced carcinogenesis and are consistently expressed in precancerous and malignant lesions [42,55].
A wide range of therapeutic vaccine platforms has been explored in preclinical and clinical studies. These include peptide-based, protein-based, DNA-based, RNA-based (including RNA replicons), viral vector-based (e.g., adenoviruses, adeno-associated viruses, alphaviruses, and vaccinia virus), bacterial vector-based (e.g., Listeria monocytogenes, Lactobacillus spp., Salmonella, Shigella, and Escherichia coli), dendritic cell-based, and whole tumor cell-based vaccines [54,56]. Many of these approaches have demonstrated promising immunogenicity and lesion regression, particularly in premalignant conditions such as cervical intraepithelial neoplasia (CIN2/3) [56].
Adoptive T-cell therapy represents another important immunotherapeutic strategy, offering precise control over the magnitude and specificity of the immune response. Clinical studies, including phase II trials in locally advanced cervical cancer (LACC), have shown encouraging therapeutic activity [57,58]. Additionally, novel vaccine designs continue to emerge. For example, electroporation-enhanced delivery of a DNA vaccine (GX-188E) targeting HPV antigens to dendritic cells has been shown to induce strong E6/E7-specific IFN-γ-producing T-cell responses in patients with CIN3 [59]. Similarly, innovative nanoparticle-based vaccines such as cPANHPVAX, which incorporate multiple HPV L2 epitopes along with E7 oncoantigens, demonstrate potential as both preventive and therapeutic agents by targeting active infections as well as established malignancies [60].
Despite these advances, therapeutic vaccines alone have shown limited efficacy in invasive cancers, and their clinical success is more evident in early-stage or premalignant lesions. Therefore, combination strategies integrating therapeutic HPV vaccines with radiotherapy, chemotherapy, immunomodulators, or immune checkpoint inhibitors are increasingly being explored to enhance treatment outcomes [56,58]. Overall, therapeutic HPV vaccination represents a promising and evolving approach in cervical cancer management, particularly when used as part of a multimodal treatment strategy.
6. Vaccine designing strategies and its application
Cancer vaccine platforms are classified into cell based vaccines, peptide-based vaccines, viral-based vaccines, and nucleic acid-based vaccines [39–41]
6.1 Peptide- and protein-based vaccines
Peptide vaccines use selected immunogenic epitopes derived from tumor or viral antigens and are attractive because of their safety, manufacturing simplicity, and personalization potential. Their main limitation is relatively low immunogenicity without adjuvants or strong delivery platforms [39,42]. Protein vaccines can improve antigen breadth, but similarly often require immunostimulatory optimization [42]. Peptide-based vaccines use short, tumor-specific antigenic peptides to stimulate immune responses. These peptides are presented by antigen-presenting cells (APCs) through HLA molecules and recognized by T cells, leading to targeted anti-tumor activity [39,43]. This platform is safe, easy to manufacture, and suitable for personalized cancer vaccines.
6.2 Cell-based vaccines
Cell-based vaccines, especially dendritic cell vaccines, exploit the potent antigen-presenting capacity of dendritic cells to activate tumor-specific T-cell responses [40,44]. Dendritic cells may be loaded with peptides, proteins, DNA, RNA, or tumor lysates ex vivo before reinfusion. Although biologically attractive, these vaccines are labor-intensive, expensive, and less scalable for broad implementation [44,45]. Cell-based vaccines include tumor cell vaccines and dendritic cell (DC) vaccines. Tumor cell vaccines use either patient-derived (autologous) or donor-derived (allogenic) tumor cells to trigger an immune response. These cells can be genetically modified to enhance their immunogenicity. However, obtaining sufficient tumor cells can be challenging [45].
Dendritic cell vaccines are more advanced, as DCs are powerful antigen-presenting cells that activate T cells. In this approach, DCs are loaded with tumor antigens such as DNA, RNA, or proteins, and have been widely tested in clinical trials with promising outcomes [44,46].
6.3 Viral vector vaccines
Viral vectors, including adenoviruses, vaccinia viruses, and alphaviruses, can induce strong innate and adaptive immune responses [41,47]. Their high immunogenicity is a key advantage, though pre-existing anti-vector immunity may reduce repeat-dose efficacy [47]. Preclinical work using adenoviral vectors encoding mutant HPV-16 E7 has shown tumor suppression and stronger antitumor responses when combined with PD-1 or PD-L1 blockade [48]. Viral vector-based vaccines utilize genetically modified viruses such as adenoviruses, poxviruses, and alphaviruses to deliver tumor antigens. These vaccines are highly immunogenic and can activate both innate and adaptive immunity [41,47]. However, pre-existing immunity against viral vectors may reduce their effectiveness. A special category, oncolytic viruses, can directly infect and kill tumor cells while also stimulating immune responses. For example, T-VEC (a modified herpes simplex virus) has shown clinical success [49].
6.4 DNA vaccines
DNA vaccines are particularly attractive in the HPV field because they are stable, relatively inexpensive, easy to manufacture, and suitable for repeated administration [18,22,50]. They can be engineered to encode E6/E7 antigens and, in some designs, to improve dendritic cell targeting or antigen presentation [50]. VGX-3100 and GX-188E are among the most widely discussed examples in the therapeutic HPV vaccine literature, with encouraging results in premalignant cervical lesions [18,22,51]. DNA vaccines remain one of the most promising therapeutic platforms for cervical precancer because of their balance of safety, antigenic precision, and translational practicality [50].
Nucleic acid-based vaccines include both DNA and mRNA vaccines. These vaccines deliver genetic material encoding tumor antigens, allowing the body to produce these antigens and stimulate immune responses. DNA vaccines are stable, cost-effective, and suitable for repeated use, but may show lower immunogenicity [50,52]. mRNA vaccines, on the other hand, provide stronger immune responses and faster development but are less stable and require special delivery systems like lipid nanoparticles [23,52]. Both platforms are promising in cervical cancer, especially those targeting HPV E6 and E7 proteins [22,51].
Figure 3: shows cancer immunotherapy
6.5 mRNA vaccines
mRNA vaccines have gained enormous attention because they can support rapid, scalable design and potent antigen expression without the need for nuclear entry [23,52,53]. Their success during the COVID-19 era has accelerated interest in oncology [53]. For cervical cancer, mRNA platforms offer the possibility of delivering E6/E7 or personalized neoantigen-encoding constructs, particularly when combined with lipid nanoparticle systems that improve stability and intracellular delivery [23,54]. Their transient expression profile may also reduce concerns associated with prolonged antigen exposure [53]. Although mRNA therapeutic vaccines in cervical cancer are still emerging, they represent an important future direction [54]. In recent years, mRNA vaccines have gained attention due to their high efficacy, safety, and rapid development potential. These vaccines work by delivering mRNA into cells, where it is translated into antigenic proteins that activate immune responses through both MHC class I and II pathways [52–54].
6.6 Nanoparticle-assisted delivery
Nanotechnology is increasingly being explored to improve delivery of drugs, genes, adjuvants, and vaccine payloads in cervical cancer [23,24,55]. Nanoparticles can enhance bioavailability, protect labile nucleic acids, support controlled release, and improve delivery to antigen-presenting cells or tumor tissue [55]. Lipid nanoparticles are especially relevant for mRNA systems, while polymeric and metal-based nanostructures are being investigated for drug delivery and theranostic purposes [23,55,56]. These platforms may also improve combination strategies by co-delivering antigenic and immunomodulatory components [55]. Nanotechnology-based delivery systems are emerging as powerful tools in cervical cancer treatment. Nanoparticles improve drug and vaccine delivery by enhancing stability, bioavailability, and targeted action [56]. Different types of nanoparticles—such as polymeric, metal-based, and carbon-based—are being explored for drug delivery, gene therapy, and immunotherapy [55,56]. Nanotechnology also enables interdisciplinary advancements by combining molecular biology, materials science, and immunology [56]. A key advancement in vaccine design is the use of adenovirus vectors, which efficiently deliver tumor antigens and induce strong immune responses. Studies have shown that vaccines like Ad-E7 (targeting HPV E7 protein) can significantly reduce tumor growth, especially when combined with immune checkpoint inhibitors (e.g., PD-1/PD-L1 blockers). This combination enhances CD8⁺ T-cell activity and reduces immunosuppressive cells, leading to better tumor control [48,57].
7. Novel Immunotherapy Combinations and Future Treatment Paradigms
Cervical cancer represents a unique model for immunotherapy due to its strong association with persistent infection by high-risk human papillomavirus (HPV), particularly HPV16 and HPV18. Viral oncoproteins E6 and E7 play a central role in carcinogenesis by inactivating key tumor suppressor pathways. Specifically, E6 promotes ubiquitin-mediated degradation of p53, impairing apoptosis and genomic stability, while E7 inactivates retinoblastoma protein (pRb), leading to uncontrolled cell cycle progression via E2F activation [20,58–60]. These oncogenic events, combined with viral antigen expression, create an immunogenic tumor environment that is highly amenable to immune-based therapeutic strategies [58,61].
• Immune Checkpoint Inhibitors and Combination Strategies
Immune checkpoint inhibitors (ICIs), particularly those targeting the PD-1/PD-L1 and CTLA-4 pathways, have revolutionized the treatment of advanced cervical cancer [49,62]. By blocking inhibitory signals that suppress T-cell activation, ICIs restore anti-tumor immune responses. While CTLA-4 primarily regulates early T-cell activation in lymphoid organs, PD-1/PD-L1 interactions dominate in peripheral tissues and the tumor microenvironment [49,63].
Recent clinical evidence (2023–2025) indicates that ICI monotherapy has limited efficacy, largely due to tumor immune evasion mechanisms [45,64]. Consequently, current research has shifted toward rational combination strategies, which aim to overcome resistance and enhance therapeutic responses [64,65]. These include:
• ICI + Chemotherapy → enhances antigen release and immunogenic cell death [66]
• ICI + Radiotherapy → promotes neoantigen exposure and T-cell infiltration [67]
• ICI + Anti-angiogenic therapy (e.g., bevacizumab) → normalizes tumor vasculature and improves immune cell access [68]
The KEYNOTE-826 and KEYNOTE-A18 trials have demonstrated that combining pembrolizumab with chemotherapy ± bevacizumab or with chemoradiotherapy significantly improves survival outcomes, establishing combination immunotherapy as a new standard of care in selected settings [44,56,69].
• Emerging Immunotherapeutic Approaches
1. Therapeutic HPV Vaccines
Unlike prophylactic vaccines, therapeutic vaccines target established infections and lesions by inducing robust cellular immune responses against E6/E7 antigens [27,32,70]. Recent studies highlight:
• Promising efficacy in CIN2/3 lesions [27,70]
• Enhanced responses when combined with ICIs [64,71]
• Development of DNA, mRNA, and viral vector platforms [50,52,72]
2. Adoptive Cell Therapy
Adoptive immunotherapy, including tumor-infiltrating lymphocytes (TILs) and engineered T-cell receptor (TCR) therapies, has shown encouraging response rates in early-phase trials [31,73]. These approaches enable the expansion and reinfusion of tumor-reactive T cells, offering personalized treatment options [73].
3. Bispecific Antibodies
Bispecific T-cell engagers (BiTEs) are an emerging class of therapeutics that simultaneously bind tumor antigens and T cells, facilitating targeted immune activation [74]. Early research suggests potential in HPV-associated malignancies [74,75].
• Tumor Microenvironment and Resistance Mechanisms
A major challenge in cervical cancer immunotherapy is the presence of an immunosuppressive tumor microenvironment (TME) [47,48,76]. Key features include:
• Regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) [76]
• Reduced antigen presentation (MHC-I downregulation) [58,76]
• Upregulation of checkpoint molecules [49,63]
• Chronic inflammation and cytokine imbalance [47]
Recent studies emphasize the need to reprogram the TME to enhance immunotherapy efficacy [76,77]. Strategies include combining ICIs with metabolic modulators, cytokine therapies, and epigenetic regulators [77]. Overall, the future of cervical cancer treatment is shifting toward precision immuno-oncology, where combination immunotherapies targeting HPV antigens, immune checkpoints, and the tumor microenvironment are expected to redefine clinical outcomes [64,71,77].
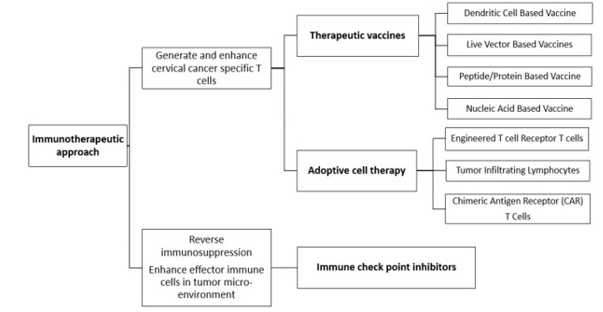
Figure 3: shows cancer immunotherapeutic approaches
8. Global challenges and implementation barriers
Despite significant scientific advancements in HPV vaccination, the global implementation of cervical cancer prevention strategies remains uneven. Several licensed HPV vaccines are available worldwide, and the World Health Organization (WHO) has continuously updated its recommendations to improve vaccine accessibility and coverage. In 2020, WHO introduced an important policy shift by endorsing a single-dose HPV vaccination schedule under appropriate conditions, based on emerging evidence showing comparable protection to two- or three-dose regimens. This strategy aims to simplify vaccine delivery, reduce costs, and improve vaccination rates, particularly in low-resource settings [48,49]. However, global vaccination coverage remains suboptimal. For example, only about 15% of girls received at least one dose of the HPV vaccine in 2021, a decline from 25% in 2019, leaving millions unprotected [48,50]. To address supply and access issues, WHO has expanded the list of prequalified vaccines. In 2024, vaccines such as Cecolin (a cost-effective bivalent vaccine from China) and Walrinvax (a two-dose vaccine) were added, helping to increase global supply and affordability [49,51]. WHO currently recommends vaccination primarily for girls aged 9–14 years, with one or two doses depending on age and health status [48,49]. The effectiveness of HPV vaccination programs also depends on implementation strategies. School-based vaccination programs have proven to be the most successful in increasing coverage, especially in low- and middle-income countries. Early vaccination before the onset of sexual activity is critical, as it not only prevents HPV infection but also contributes to herd immunity [52,53]. Despite these efforts, multiple barriers continue to hinder widespread vaccine uptake. In regions such as Sub-Saharan Africa, challenges include limited healthcare infrastructure, funding constraints, cultural beliefs, and logistical difficulties. Similarly, even in high-income countries like the United States, vaccine hesitancy, misinformation, and socio-economic disparities affect vaccination rates. In the Middle East and North Africa, the lack of organized screening programs further complicates cervical cancer control [50,54]. Key global challenges include high vaccine costs, limited supply, weak adolescent immunization systems, misinformation, cultural resistance, inadequate screening programs, and poor access to treatment facilities. Addressing these barriers requires not only scientific innovation but also strong public health policies, community awareness, and healthcare system strengthening [48,52,54]. For countries such as India, additional priorities include increasing public awareness about HPV vaccination, integrating vaccination into existing reproductive health programs, expanding cervical cancer screening coverage, and ensuring timely access to treatment for diagnosed cases. Ultimately, while HPV vaccines offer a powerful tool for prevention, the elimination of cervical cancer will depend on equitable and effective implementation strategies worldwide [49,53].
Future directions
For combating HPV infection in the human population, there is a need to strengthen awareness, social counselling, and clinical assistance, particularly among young women. In addition, accurate and high-quality diagnostic approaches are essential for improving therapeutic outcomes and early disease detection [48,55]. As research continues to advance, greater emphasis is being placed on the development of more efficient vaccines to control HPV-associated pathogenesis.
There is a growing need to integrate advanced immunotherapeutic strategies with vaccination approaches. In particular, mRNA-based vaccines with high therapeutic efficacy are emerging as promising tools due to their rapid development potential and strong immunogenicity [23,56]. Personalizing vaccines based on tumor-specific mutations and neoantigen profiling may further enhance therapeutic efficacy [58,59]. Combination strategies integrating therapeutic vaccines with immune checkpoint inhibitors (ICIs) or standard therapies such as chemotherapy and radiotherapy are expected to boost anti-tumor responses and improve clinical outcomes [45,60]. Additionally, the application of bioinformatics tools for epitope prediction and vaccine optimization is becoming increasingly important in next-generation vaccine design [58]. For effective long-term control, increasing vaccination coverage, strengthening screening programs, and ensuring equitable access to treatment by 2030 remain critical goals aligned with WHO elimination strategies [48,49]. Future research should prioritize biomarker-driven precision immunotherapy. Several directions appear particularly important for the next phase of cervical cancer therapeutics. First, therapeutic vaccine design is likely to move toward more potent, multi-antigen, and personalized platforms, including neoantigen-informed and bioinformatics-guided strategies [58,59]. Second, mRNA and other nucleic acid technologies are expected to expand due to their adaptability and compatibility with nanoparticle-based delivery systems [23,56]. Third, rational combinations with immune checkpoint inhibitors, radiotherapy, and standard chemotherapeutics may improve response durability and overcome resistance [45,60]. Fourth, biomarker-guided management, including circulating cell-free DNA (cfDNA), tumor mutational burden (TMB), and immune monitoring approaches, may help refine prognosis and treatment selection [25,61]. Finally, public health progress toward elimination will require convergence of biomedical innovation with implementation science, affordability, and policy support [48,49].
Conclusion
Cervical cancer remains a significant global health burden, but the advent of prophylactic and therapeutic vaccines offers unprecedented promise in its prevention and treatment. While current vaccines have demonstrated efficacy in reducing HPV-related disease incidence, efforts must now focus on overcoming barriers to access, improving therapeutic vaccine performance, and harnessing novel technologies [48,49]. The integration of immunology, molecular biology, and digital tools will be central to shaping the future landscape of cervical cancer therapeutics. A comprehensive, equitable approach combining vaccination, screening, and treatment is essential for achieving long-term eradication of cervical cancer [23,56]. Cervical cancer remains a leading but largely preventable malignancy among women, especially in resource-limited settings. Persistent high-risk HPV (hrHPV) infection drives most cases, and improved understanding of HPV biology, immune evasion, and tumor–host interactions has opened new therapeutic opportunities [4,6]. Prophylactic vaccines remain foundational for prevention, but therapeutic vaccines are increasingly important for persistent infection, precancerous lesions, and potentially invasive disease [4,45]. Advances in DNA vaccines, mRNA technologies, viral vectors, dendritic cell approaches, and nanoparticle-based delivery systems are reshaping the future of cervical cancer therapeutics [23,22,56]. At the same time, immune checkpoint inhibitors and rational combination immunotherapy strategies are expanding treatment possibilities for advanced disease [45,60]. The future of cervical cancer control will depend not only on improved vaccine design and translational immunology, but also on equitable access to vaccination, effective screening programs, and timely treatment [48,49]. A coordinated multidisciplinary strategy integrating biomedical innovation with public health implementation remains essential for achieving sustained disease reduction and eventual global elimination of cervical cancer [49].
Abbreviations
• APC – Antigen-Presenting Cell
• BiTE – Bispecific T-cell Engager
• CAR-T – Chimeric Antigen Receptor T-cell
• CC – Cervical Cancer
• cfDNA – Circulating Cell-Free DNA
• CIN – Cervical Intraepithelial Neoplasia
• CTL – Cytotoxic T Lymphocyte
• DC – Dendritic Cell
• DNA – Deoxyribonucleic Acid
• dsDNA – Double-Stranded DNA
• E6/E7 – HPV Oncoproteins E6 and E7
• E2F – E2 Promoter Binding Factor
• E6-AP – E6-Associated Protein
• FDA – Food and Drug Administration
• FIGO – International Federation of Gynecology and Obstetrics
• GX-188E – HPV Therapeutic DNA Vaccine
• HPV – Human Papillomavirus
• hrHPV – High-Risk Human Papillomavirus
• HIV – Human Immunodeficiency Virus
• HLA – Human Leukocyte Antigen
• ICI – Immune Checkpoint Inhibitor
• IFN-γ – Interferon Gamma
• IgG – Immunoglobulin G
• L1/L2 – HPV Capsid Proteins
• LACC – Locally Advanced Cervical Cancer
• LCR – Long Control Region
• LMICs – Low- and Middle-Income Countries
• MAPK – Mitogen-Activated Protein Kinase
• MDSCs – Myeloid-Derived Suppressor Cells
• MHC-I – Major Histocompatibility Complex Class I
• mRNA – Messenger RNA
• MSI-H – Microsatellite Instability-High
• NAb – Neutralizing Antibody
• NK cells – Natural Killer Cells
• PD-1 – Programmed Cell Death Protein 1
• PD-L1 – Programmed Death-Ligand 1
• PI3K/Akt/mTOR – Phosphoinositide 3-Kinase / Protein Kinase B / Mammalian Target of Rapamycin
• p53 – Tumor Protein 53
• pRb – Retinoblastoma Protein
• TCR – T-cell Receptor
• TILs – Tumor-Infiltrating Lymphocytes
• TMB – Tumor Mutational Burden
• TME – Tumor Microenvironment
• Tregs – Regulatory T Cells
• VLPs – Virus-Like Particles
• WHO – World Health Organization
References
- Doorbar J, Egawa N, Griffin H, Kranjec C, Murakami I. (2015). Human papillomavirus molecular biology and disease association. Rev Med Virol. 25(S1):2–23.
View at Publisher | View at Google Scholar - Graham SV. (2017). The human papillomavirus replication cycle. Nat Rev Microbiol. 15(11):684–698.
View at Publisher | View at Google Scholar - Crosbie EJ, Einstein MH, Franceschi S, Kitchener HC. (2013). Human papillomavirus and cervical cancer. Lancet. 382(9895):889–899.
View at Publisher | View at Google Scholar - Trimble CL, Morrow MP, Kraynyak KA, et al. (2015). Safety, efficacy, and immunogenicity of VGX-3100. Lancet. 386(10008):2078–2088.
View at Publisher | View at Google Scholar - Tewari KS, Monk BJ. (2019). New strategies in cervical cancer. Lancet Oncol. 20(1):e26–e35.
View at Publisher | View at Google Scholar - Sung H, Ferlay J, Siegel RL, et al. (2023). Global cancer statistics 2022. CA Cancer J Clin. 73(1):17–48.
View at Publisher | View at Google Scholar - Arbyn M, Weiderpass E, Bruni L, et al. Estimates of incidence and mortality. Lancet Glob Health. 2020;8(2):e191–e203.
View at Publisher | View at Google Scholar - Schiffman M, Castle PE. (2007). HPV: epidemiology and prevention. Nat Rev Cancer. 7(1):11–22.
View at Publisher | View at Google Scholar - Plummer M, Peto J, Franceschi S. (2012). Smoking and cervical cancer. Int J Cancer. 130(1):1–9.
View at Publisher | View at Google Scholar - Mitra A, MacIntyre DA, Lee YS, et al. (2015). Cervical microbiota and HPV persistence. Sci Transl Med. 7(293):293ra94.
View at Publisher | View at Google Scholar - (2020). WHO. Global strategy to accelerate elimination of cervical cancer.
View at Publisher | View at Google Scholar - Bruni L, Albero G, Serrano B, et al. (2023). ICO/IARC HPV information centre report.
View at Publisher | View at Google Scholar - Clifford GM, Smith JS, Plummer M, et al. (2003). HPV types in cervical cancer. Lancet. 362:991–998.
View at Publisher | View at Google Scholar - Doorbar J. (2005). The papillomavirus life cycle. J Clin Virol. 32:S7–S15.
View at Publisher | View at Google Scholar - McBride AA. (2012). Oncogenic HPV integration. Nat Rev Cancer. 12(7):497–507.
View at Publisher | View at Google Scholar - Scheffner M, Werness BA, Huibregtse JM, et al. (1990). HPV E6 targets p53. Cell. 63(6):1129–1136.
View at Publisher | View at Google Scholar - Dyson N, Howley PM, Münger K, Harlow E. (1989). E7 and Rb interaction. Science. 243:934–937.
View at Publisher | View at Google Scholar - Moody CA, Laimins LA. (2010). HPV pathogenesis. Nat Rev Cancer. 10(8):550–560.
View at Publisher | View at Google Scholar - Stanley M. (2006). Immune responses to HPV. Vaccine. 24:S16–S22.
View at Publisher | View at Google Scholar - zur Hausen H. (2002). Papillomaviruses in cancer. Nat Rev Cancer. 2(5):342–350.
View at Publisher | View at Google Scholar - Mo Y, Ma J, Zhang H, et al. (2022). HPV vaccines: current scenario. Front Cell Infect Microbiol. 12:909223.
View at Publisher | View at Google Scholar - Harper DM, Franco EL, Wheeler C, et al. (2004). HPV vaccine efficacy. Lancet. 364:1757–1765.
View at Publisher | View at Google Scholar - Garland SM, Hernandez-Avila M, Wheeler CM, et al. (2007). Quadrivalent HPV vaccine. N Engl J Med. 356:1928–1943.
View at Publisher | View at Google Scholar - Joura EA, Giuliano AR, Iversen OE, et al. (2015). 9-valent HPV vaccine. N Engl J Med. 372:711–723.
View at Publisher | View at Google Scholar - (2024). WHO. Weekly epidemiological record HPV vaccines.
View at Publisher | View at Google Scholar - Schiller JT, Lowy DR. (2012). HPV vaccine development. Nat Rev Microbiol. 10(10):681–692.
View at Publisher | View at Google Scholar - Trimble CL, et al. (2015). VGX-3100 trial. Lancet. 386:2078–2088.
View at Publisher | View at Google Scholar - Kim TJ, Jin HT, Hur SY, et al. (2014). GX-188E DNA vaccine. Clin Cancer Res. 20:161–172.
View at Publisher | View at Google Scholar - Melief CJM, van Hall T, Arens R, et al. (2015). Therapeutic cancer vaccines. J Clin Invest. 125(9):3401–3412.
View at Publisher | View at Google Scholar - van Poelgeest MIE, Welters MJP, van Esch EMG, et al. (2013). HPV vaccine clinical responses. J Clin Oncol. 31:197–205.
View at Publisher | View at Google Scholar - Stevanović S, Draper LM, Langhan MM, et al. (2015). TIL therapy in cervical cancer. J Clin Oncol. 33:1543–1550.
View at Publisher | View at Google Scholar - Rosales R, Lopez-Contreras M, Rosales C, et al. (2017). HPV therapeutic vaccines. Expert Rev Vaccines. 16:1227–1240.
View at Publisher | View at Google Scholar - Butterfield LH. (2015). Cancer vaccines. BMJ. 350:h988.
View at Publisher | View at Google Scholar - Palucka K, Banchereau J. (2012). Dendritic cell-based vaccines. Nat Rev Cancer. 12:265–277.
View at Publisher | View at Google Scholar - Draper SJ, Heeney JL. Viral vectors. (2010). Nat Rev Microbiol. 8:62–73.
View at Publisher | View at Google Scholar - Kaufman HL, Kohlhapp FJ, Zloza A. (2015). Oncolytic viruses. Nat Rev Drug Discov. 14:642–662.
View at Publisher | View at Google Scholar - Kim YC, Jarrahian C, Zehrung D, et al. (2021). DNA vaccines. NPJ Vaccines. 6:13.
View at Publisher | View at Google Scholar - Pardi N, Hogan MJ, Porter FW, Weissman D. (2018). mRNA vaccines. Nat Rev Drug Discov. 17:261–279.
View at Publisher | View at Google Scholar - Sahin U, Karikó K, Türeci Ö. (2014). mRNA cancer vaccines. Nat Rev Drug Discov. 13:759–780.
View at Publisher | View at Google Scholar - Hou X, Zaks T, Langer R, Dong Y. (2021). Lipid nanoparticles. Nat Rev Mater. 6:1078–1094.
View at Publisher | View at Google Scholar - Peer D, Karp JM, Hong S, et al. (2007). Nanocarriers. Nat Nanotechnol. 2:751–760.
View at Publisher | View at Google Scholar - Shi J, Kantoff PW, Wooster R, Farokhzad OC. (2017). Nanomedicine. Nat Rev Cancer. 17:20–37.
View at Publisher | View at Google Scholar - Burtness B, Harrington KJ, Greil R, et al. (2019). Pembrolizumab trials. Lancet. 394:1915–1928.
View at Publisher | View at Google Scholar - Colombo N, Dubot C, Lorusso D, et al. (2021). KEYNOTE-826. N Engl J Med. 385:1856–1867.
View at Publisher | View at Google Scholar - Lorusso D, Colombo N. (2022). Immunotherapy cervical cancer. Ann Oncol. 33:561–572.
View at Publisher | View at Google Scholar - Naumann RW, Hollebecque A, Meyer T, et al. (2019). Pembrolizumab cervical cancer. J Clin Oncol. 37:1470–1478.
View at Publisher | View at Google Scholar - Hinshaw DC, Shevde LA. (2019). Tumor microenvironment. Cancer Res. 79:4557–4566.
View at Publisher | View at Google Scholar - Binnewies M, Roberts EW, Kersten K, et al. (2018). Tumor immune landscape. Nat Med. 24:541–550.
View at Publisher | View at Google Scholar - Sharma P, Allison JP. (2015). Immune checkpoint therapy. Cell. 161:205–214.
View at Publisher | View at Google Scholar - Topalian SL, Drake CG, Pardoll DM. (2015). Immune checkpoint blockade. Cancer Cell. 27:450–461.
View at Publisher | View at Google Scholar - Monk BJ, Tewari KS. (2024). Advances in cervical cancer therapy. Lancet Oncol.
View at Publisher | View at Google Scholar - Cohen EEW, Soulières D, Le Tourneau C, et al. (2023). Immunotherapy advances. J Clin Oncol.
View at Publisher | View at Google Scholar - (2024). NCI. Cervical cancer treatment updates.
View at Publisher | View at Google Scholar - (2023). WHO. Cervical cancer elimination progress report.
View at Publisher | View at Google Scholar - (2024). FDA. Pembrolizumab approval cervical cancer.
View at Publisher | View at Google Scholar - (2024). ClinicalTrials.gov. KEYNOTE-A18 trial data.
View at Publisher | View at Google Scholar - (2023–2025). Recent PubMed reviews on HPV immunotherapy combinations.
View at Publisher | View at Google Scholar - Moody CA, Laimins LA. (2010). Human papillomavirus oncoproteins: pathways to transformation. Nat Rev Cancer. 10(8):550–560.
View at Publisher | View at Google Scholar - McLaughlin-Drubin ME, Münger K. (2009). Oncogenic activities of HPV E6 and E7. Virology. 384(2):335–344.
View at Publisher | View at Google Scholar - Mantovani F, Banks L. (2001). The human papillomavirus E6 protein and its contribution to malignancy. Semin Cancer Biol. 11(6):485–493.
View at Publisher | View at Google Scholar - Stanley M. (2006). Immune responses to human papillomavirus. Vaccine. 24(S3):S16–S22.
View at Publisher | View at Google Scholar - Chung HC, Ros W, Delord JP, et al. (2019). Pembrolizumab in advanced cervical cancer (KEYNOTE-158). J Clin Oncol. 37(17):1470–1478.
View at Publisher | View at Google Scholar - Pardoll DM. (2012). The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 12(4):252–264.
View at Publisher | View at Google Scholar - Lorusso D, Colombo N. (2022). Immunotherapy in cervical cancer: current status and future perspectives. Ann Oncol. 33(6):561–572.
View at Publisher | View at Google Scholar - Frenel JS, Le Tourneau C, O'Neil B, et al. (2017). Safety and efficacy of immune checkpoint inhibitors. J Clin Oncol. 35(36):4035–4041.
View at Publisher | View at Google Scholar - Galluzzi L, Buqué A, Kepp O, Zitvogel L, Kroemer G. (2017). Immunogenic cell death in cancer therapy. Nat Rev Immunol. 17(2):97–111.
View at Publisher | View at Google Scholar - Demaria S, Golden EB, Formenti SC. (2015). Radiotherapy and immunotherapy synergy. Nat Rev Clin Oncol. 12(12):731–741.
View at Publisher | View at Google Scholar - Hegde PS, Wallin JJ, Mancao C. (2016). Predictive markers and combination strategies. Cancer Cell. 29(6):821–832.
View at Publisher | View at Google Scholar - Colombo N, Dubot C, Lorusso D, et al. (2021). Pembrolizumab + chemotherapy in cervical cancer (KEYNOTE-826). N Engl J Med. 385(20):1856–1867.
View at Publisher | View at Google Scholar - Trimble CL, et al. (2015). Therapeutic HPV vaccine VGX-3100 efficacy. Lancet. 386(10008):2078–2088.
View at Publisher | View at Google Scholar - Melief CJM, van Hall T, Arens R, et al. (2015). Therapeutic cancer vaccines. J Clin Invest. 125(9):3401–3412.
View at Publisher | View at Google Scholar - Pardi N, Hogan MJ, Porter FW, Weissman D. (2018). mRNA vaccines: a new era. Nat Rev Drug Discov. 17(4):261–279.
View at Publisher | View at Google Scholar - Stevanović S, Draper LM, Langhan MM, et al. (2015). Adoptive T-cell therapy for cervical cancer. J Clin Oncol. 33(14):1543–1550.
View at Publisher | View at Google Scholar - Labrijn AF, Janmaat ML, Reichert JM, Parren PWHI. (2019). Bispecific antibodies: a review. Nat Rev Drug Discov. 18(8):585–608.
View at Publisher | View at Google Scholar - Goebeler ME, Bargou RC. (2020). T cell–engaging therapies (BiTEs). Nat Rev Clin Oncol. 17(7):418–434.
View at Publisher | View at Google Scholar - Binnewies M, Roberts EW, Kersten K, et al. (2018). Tumor immune microenvironment. Nat Med. 24(5):541–550.
View at Publisher | View at Google Scholar - Hinshaw DC, Shevde LA. (2019). Tumor microenvironment and immunotherapy. Cancer Res. 79(18):4557–4566.
View at Publisher | View at Google Scholar
